|
|
 |
|
О родовых разграничениях лепидодендровых карбона
Автор: donbassfossil
Рубрика: Палеонтологические публикации
Приступая к любительским палеонтологическим изысканиям карбоновых растений, за последние три года я прошел нелегкий путь от полного непонимания предмета изысканий (когда образец молодой коры лепидодендрона принимается за «змеиную кожу») до попыток систематизировать найденные ископаемые растения в группы в соответствии с существующим порядком классификации (различать роды плауновидных, оперируя при этом терминами кладограмма, плезиоморфный признак или что-то еще в духе филогенетических наворотов). Нет, конечно, я далек от иллюзии, что за это время превратился в профессионального палеонтолога, однако, получил ощущение, которое знакомо тем, кто осваивая английский, в какой-то момент почувствовали, что, при чтении книги в оригинале, они уже способны следить за сюжетом произведения, а не просто мучительно складывать слова. То есть, совсем даже не англичанин, но уже и не «отскакивает как от стенки».
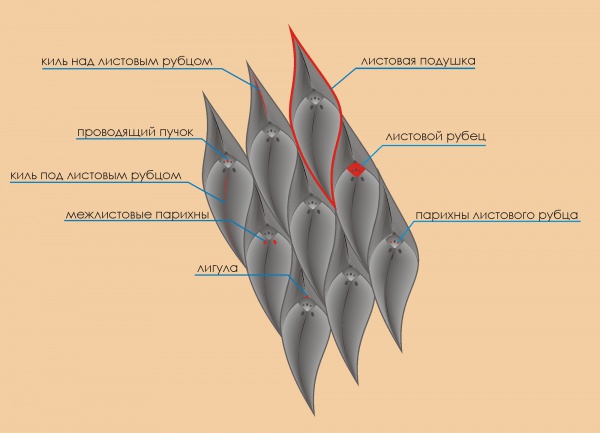
Одним из наиболее часто встречающихся на «охоте» родов является лепидодендрон, относящийся к плауновидным. Для лепидодендронов же характерно огромное количество имен видов, которые щедро раздавали палеонтологи прошлого, к ужасу начинающих выстроив сложнейшую систему номенклатуры этих растений. Ознакомившись с доступными источниками в Интернет, такими как коллекция Kidston или систематизация Лангфорда, я понял, что существуют десятки видов найденных растений, и главным критерием внутривидового разделения является морфология листовой подушки коры отпечатков лепидодендрона. Ее форма, размер, пропорции, наличие или отсутствие некоторых деталей на листовом рубце или листовой подушке. Здесь следует сказать, что находимые трехмерные образцы ископаемых растений встречаются в двух основных формах: отпечатки (только форма) и анатомической сохранности (когда внутренне строение, скажем, ствола сохранено, при этом органика полностью замещена минералом). В общем, я несколько упрощенно изложил формы фоссилизации растений, памятуя о том, что уже писал об этом в прежней публикации. Но суть в данном случае заключается не в деталях, а в том, что анатомическая сохранность встречается очень редко. За три года, просмотрев тысячи образцов, я нашел единственный маленький со следами сохранившейся клеточной структуры. Именно по этой причине в прошлом в основу классификации лепидодендронов была положена морфология листовых подушек, а не анатомические особенности растений.
Ниже я привожу перевод фрагмента публикации американского палеонтолога ДиМишеля, который является своего рода «взрывателем» традиций систематики лепидодендронов, взяв за основу как раз анатомию, а не морфологию растения. Это не цитата, перевод вольный, но взят в кавычки, чтобы было ясно, где мысли американца, а где мои. Думаю, что этот фрагмент будет полезен тем, кто интересуется разнообразием плауновидных ископаемых.
«Выстраивание ископаемых растений в соответствии с их таксонометрической иерархией, которая бы отражала филогенетические связи или, по крайней мере, даже их общие морфологические характеристики всегда было проблематичной частью палеоботаники. Трудности в установлении естественных связей между исчезнувшими растениями сосредоточены вокруг недостаточности количества образцов, где была бы органическая связь между вегетативными и репродуктивными частями растения, недооценка значимости особенностей морфологии развития в сравнении с методами оценки статических образцов и великое разнообразие имен, присвоенных образцам одного вида с различной степенью сохранности. Эта оценка полной мерой относится к лепидодендровым карбона.
Годы предшествующих исследований принесли изобилие морфологической информации, но также и сформировали запутанную номенклатурную сеть, которая сдерживала распознавание конкретных форм и эволюционных связей между ними. Причины сложившейся ситуации достаточно сложны, уходят корнями в традиционные подходы к определению конкретных форм и искренней уверенности в том, что анатомия растений не является ценным диагностическим инструментом и не коррелирует с характерной (уникальной) формой листовой подушки, которая служила основой определения большинства родов ископаемых растений. Самой существенной из всех проблемой было то, что имя Lepidodendron было необоснованно распространено как на компрессионные формы (отпечатки), так и на образцы окаменевшей древесины.
Типичным представителем рода Lepidodendron является L. aculeatum Sternb., который базировался на отпечатках с характерными вертикально удлиненными листовыми подушками, имевшими межлистовые парихны.
Единственным видом, для которого был найден ствол анатомической сохранности да еще и с листовыми подушками характерной формы типа L. aculeatum, имеющими межлистовые парихны, был L. hickii Watson (1907). Другие формы с анатомической сохранностью были помещены в Лепидодендроны, полностью или частями, и представляли собой самом деле Paralycopodites brevifoliusi (DiMichele 1980), Lepidophloios (DiMichele 1979a), Sublepidophloios и группу лепидодендридов, включающую "Lepidodendron" vasculare (DiMichele 1981).
Эти уникальные вегетативные формы, отличаясь друг от друга архитектурой листовых подушек и анатомией, коррелировали с различными репродуктивными структурами, что лишь усиливало их уникальность. Применение конкретных имен имеет вторичное по важности значение для осознания того, что род является уникальным, и не все они являются лепидодендронами, несмотря на сложившуюся многолетнюю практику такой трактовки.
Как уже говорилось, анатомически сохраненный Lepidodendron hickii максимально приближается по свойствам к L. aculeatum. Характерные особенности анатомии L. hickii сравнительно с другими лепидодендридами, у которых соотношение высота/ширина листовой подушки более 1, позволили сделать вывод, что такое соотношение не является базисом для родового разграничения, что оно, по сути, является плезиоморфным признаком и, таким образом, теряет свою значимость.
Другой важной группой плауновидных, для которых характерна «лепидодендроновая» форма листовых подушек (H/W более 1), не меняющая пропорции на любых стадиях развития растения (т.е. независимо от диаметра ствола или ветви) является "Lepidodendron" vasculare и его сородичи. Кроме того, на листовых пдушках отсутствуют межлистовые парихны и они располагаются на стволах растений, отличающихся от L. hickii как анатомически, так и морфологией репродуктивных органов (в дальнейшем, автор даст ему новое родовое имя Diaphorodendron). Следует сказать, что Lepidophloios также может иметь вертикально удлиненные листовые подушки на определенных стадиях развития.
Выявление более, чем одного анатомического и репродуктивного различий форм с вертикально удлиненными листовыми подушками позволяет использовать наличие или отсутствие межлистовых парихн как признак для разграничения отпечатков, даже не имея анатомических данных.
Структурно сохранившиеся древовидные плауновые с листовыми подушками подобными Lepidodendron aculeatum были подобраны последовательно, чтобы получить внутреннюю структуру, связанную с L. hickii. Это были образцы периода Westphalian A из Англии (Watson 1907), Донецкого бассейна (Zalessky 1909, 1911), Нидерландов (Koopmans 1928,1934), США (бассейн Иллинойса).
Единственно несовпадение с Lepidodendron aculeatum имели три образца, имевших листовые подушки как у L. aculeatum, но анатомию как у Lepidophloios, в частности коронированная сифоностела. Эти образцы были описаны Сьюардом (Seward 1906) и Залесским как Lepidodendron aculeatum и Скоттом (Scott 1906) как L. obovatum. Позже, в 1970 году, Томас (Thomas) предложил считать L. aculeatum и L. obovatum синонимами. Образец Сьюарда был главным аргументом в пользу утвердившегося мнения о том, что анатомия может не учитываться при разграничении родов Lepidodendron и Lepidophloios и как следствие, что эти два рода имели постепенно преходящую друг к другу морфологию. Образец Залесского имел листовые подушки в форме лепидодендрона, но частично перекрывающимися, как подушки у Sublepidophloios Sterzel (Hopping 1956). Sublepidophloios отличается от Lepidophloios тем, что листовые рубцы расположены как у лепидофлойоза, а форма листовой подушки как у лепидодендрона.
Необходимо сказать, что у родов Asolanus и Bothrodendron все еще недостает образцов анатомической сохранности, чтобы четче понять, куда их следует отнести».
Подробнее о других плауновидных я писал в этой публикации, а также в этой.
Источник неофициального перевода: William A. DiMichele, Lepidodendron hickii and Generic Delimitation in Carboniferous Lepidodendrid Lycopods, Systematic Botany, Vol. 8, No. 3 (Jul. - Sep., 1983), 317-333.
Глоссарий
Плезиоморфные признаки - предковые признаки, термин, предложенный У. Хеннингом (1950) для обозначения примитивных признаков. Э. Майр (1969) использовал его в качестве синонима термина “предковый признак” (ancestral). Признак является предковым, если он не изменился существенным образом по сравнению с гомологичным признаком у предка, и является производным, если он существенно изменился. Например, наличие крыльев является предковым признаком у нелетающих птиц, но производным — у птиц в целом по сравнению с рептилиями.
Апоморфный признак - признак, характеризирующий (в отличие от плезиоморфного) только данную таксономическую группу, эволюционирующий в ее пределах и полностью отсутствующий в других группах.
Ключевые слова (тэги): карбон, лепидодендрон, Lepidodendron
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 6 февраля 2012 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://ammonit.ru./text/364.htm#57820">http://ammonit.ru./text/364.htm#57820</a> |
|
|||||
Интересная у Вас получилась публикация! | ||||||
|
комментарий 1 уровня |
Комментарий создан 6 февраля 2012 года в 23:55:54 |
|||||
 Добавить в избранное
Добавить в избранное 


|
<a href="http://ammonit.ru./text/364.htm#57821">http://ammonit.ru./text/364.htm#57821</a> |
|
|||||
Рад, что Вам понравилось. | ||||||
|
комментарий 2 уровня |
Комментарий создан 7 февраля 2012 года в 00:03:04 |
|||||

|
<a href="http://ammonit.ru./text/364.htm#57832">http://ammonit.ru./text/364.htm#57832</a> |
|
|||||
Прочитал, большое спасибо. Помогает понять. Хотя еще очень много незнакомого. | ||||||
|
комментарий 1 уровня |
Комментарий создан 7 февраля 2012 года в 00:28:55 |
|||||
|
<a href="http://ammonit.ru./text/364.htm#57875">http://ammonit.ru./text/364.htm#57875</a> |
|
|||||
Для меня самого, как и для Вас, еще очень много незнакомого. Работает принцип - чем дальше в карбоновый лес, тем больше "лепидодендров". | ||||||
|
комментарий 2 уровня |
Комментарий создан 7 февраля 2012 года в 09:37:35 |
|||||
|
<a href="http://ammonit.ru./text/364.htm#57972">http://ammonit.ru./text/364.htm#57972</a> |
|
|||||
Для начала перечитываю Ваши публикации и публикации Дмитрия. Когда возвращаешься к ним через некоторое время. знакомых слов и понятий становится больше. По Вашим следам идти легче. Получаются хорошие "выжимки" по конкретным вопросам. | ||||||
|
комментарий 3 уровня |
Комментарий создан 7 февраля 2012 года в 21:24:03 |
|||||
|
<a href="http://ammonit.ru./text/364.htm#58032">http://ammonit.ru./text/364.htm#58032</a> |
|
|||||
Спасибо. Это не альтруизм, а прагматика. В соответствии с Пирамидой обучения:
| ||||||
|
комментарий 4 уровня |
Комментарий создан 7 февраля 2012 года в 23:23:47 |
|||||
|
<a href="http://ammonit.ru./text/364.htm#58114">http://ammonit.ru./text/364.htm#58114</a> |
|
|||||
Интересная пирамида. | ||||||
|
комментарий 5 уровня |
Комментарий создан 8 февраля 2012 года в 12:29:07 |
|||||
|
<a href="http://ammonit.ru./text/364.htm#57902">http://ammonit.ru./text/364.htm#57902</a> |
|
|||||
Хоть и не моя тема, а интересно было. Спасибо.
| ||||||
|
комментарий 1 уровня |
Комментарий создан 7 февраля 2012 года в 12:05:46 |
|||||

{kind=link}
{kind=link}
|
<a href="http://ammonit.ru./text/364.htm#57927">http://ammonit.ru./text/364.htm#57927</a> |
|
|||||
Спасибо, загадки и возможные разгадки - вот что подогревает интерес. | ||||||
|
комментарий 2 уровня |
Комментарий создан 7 февраля 2012 года в 17:10:23 |
|||||