|
|
 |
|
Пару слов о лепидодендронах
Автор: donbassfossil
Рубрика: Палеонтологические публикации
Лепидодендрон – один из многих давно и безвозвратно исчезнувших родов древовидных растений, и является, пожалуй, самым характерным представителем флоры карбоновых болот. Легко запоминающимся он является по причине весьма своеобразной формы коры, напоминающей чешую. Чешуйчатое дерево – второе название лепидодендрона. О лепидодендронах известно многое, однако я не встречал однозначно четкого и понятного изложения материала для неспециалиста. Впрочем, возможно, пока еще мало читал. Самое основное это то, что классическая систематизация и, соответственно, видовые различия, основаны на форме и пропорциях листовых подушек (ЛП) и листового рубца (ЛР), местоположением ЛР на ЛП, наличии/отсутствии межлистовых парихн и т.д. В последнее время, правда, стали вноситься признаки видового различия, основанные на форме клеток кутикулы, наличии устьиц и др. тонкостях клеточной структуры эпидермиса. Схематически кора выглядит следующим образом.
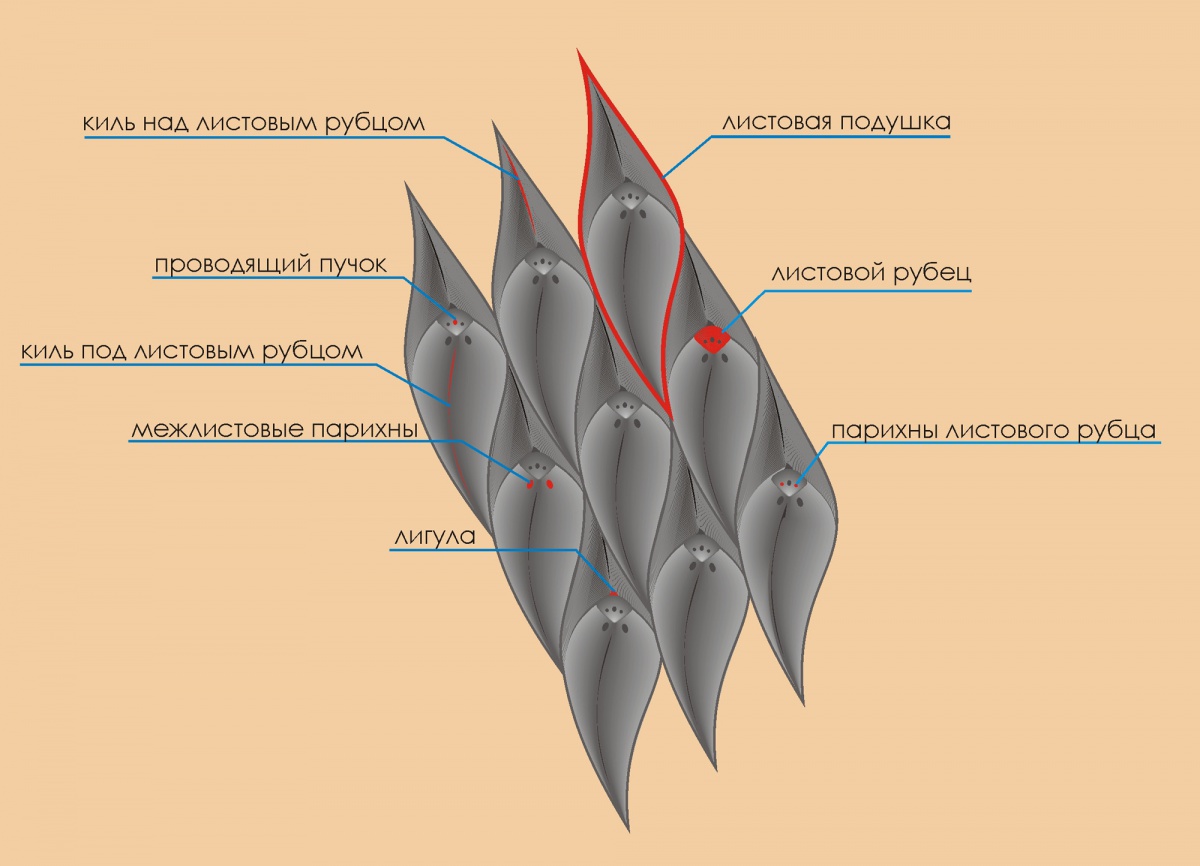
Напоминаю, что парихны были ответственны за газовый обмен (углекислота и кислород), проводящий пучок обеспечивал «водоснабжение» листа.
Кора реальных образцов выглядит так (здесь показаны образцы L. aculeatum из коллекции автора с плотно подогнанными листовыми подушками).


Ромбические чешуи на коре лепидодендрона располагаются либо плотно прилегая друг к другу, либо на некотором расстоянии, причем промежутки, как правило, заполнены морщинистыми складками коры, что указывает на вторичный рост стебля после возникновения листовых подушек и некоторые деформации поверхности.
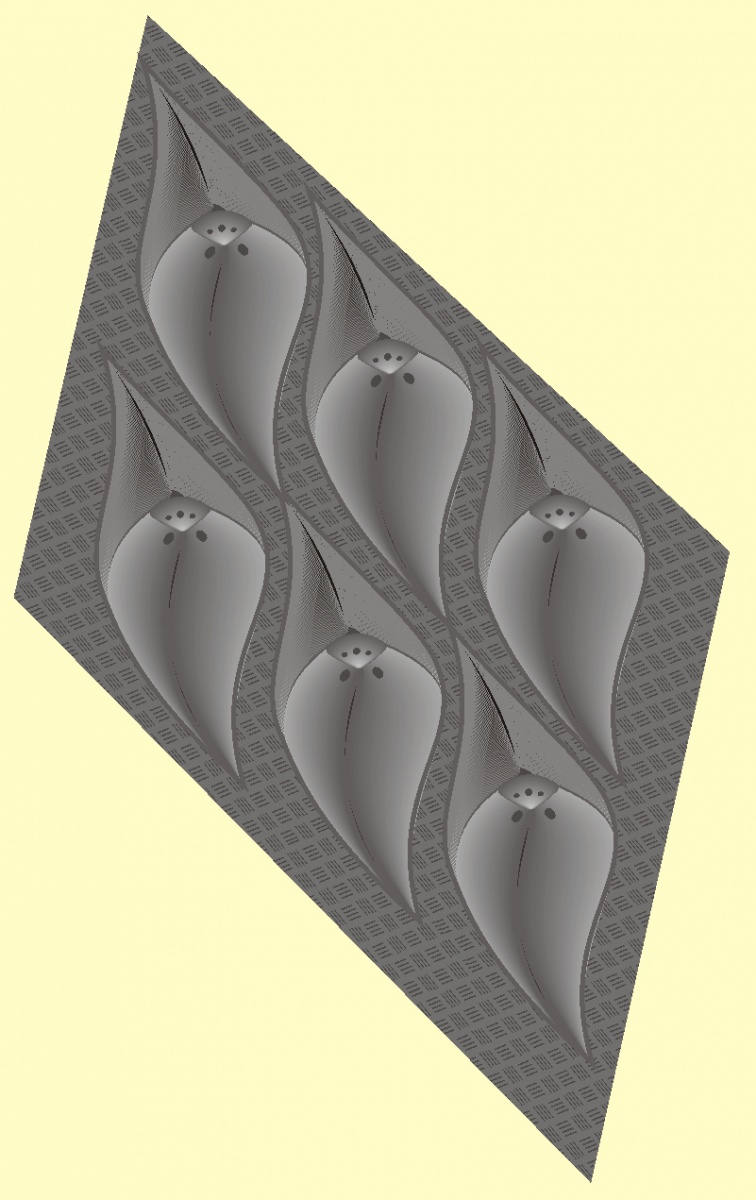
Причем, например, Lepidodendron mannebachense практически не имел зазоров коры между листовыми подушками и не имел вторичного роста,

а L. serpentigerum, L. aculeatum или L. veltheimi, напротив демонстрируют способность ветвей и ствола расширяться по мере развития и роста, причем деформации наблюдаются не только на листовых подушках, но иногда и на листовых рубцах.

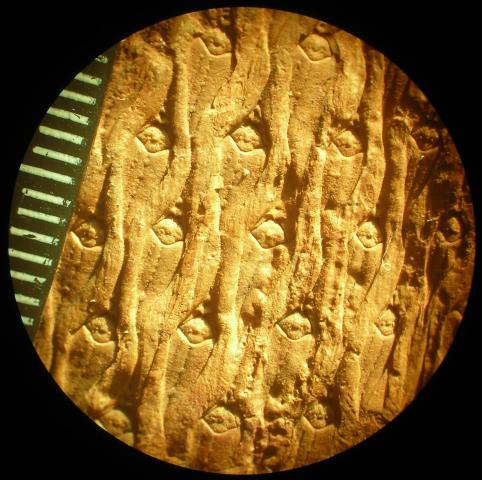
Я намеренно не буду останавливаться на изложении общеизвестных фактов о том, что лепидодендроны являлись плауновидными древовидными растениями, имели высоту ствола 30-35 метров, диаметр ствола до 2м и буйно процветали на Земле с конца девона до начала перми.
Схематическое изображение и некоторые образцы моих находок можно просмотреть здесь .
Коротко остановлюсь на некоторых проблемных вопросах систематики и вопросах онтогенеза.
Что касается систематизации, то я всегда испытывал и по сей день испытываю проблемы с определением видовой принадлежности лепидодендронов. То есть, наличие огромного количества видов типа L. aculeatum, L. bretonense, L. brevifolium, L. calamopsoides, L. carinatum, L. chemungense, L. clypeatum, L. corrugatum, L. dicentricum, L. dichotomum, L. elegans, L. epididymis, L. esnostense, L. gaspianum, L. giganteum, L. gracile, L. hangingeri, L. harcourtii, L. hickii, L. jaraczewskii, L. johnsonii, L. lanceolatum, L. lineare, L. loricatum, L. lycopodioides, L. modulatum, L. obovatum, L. ophiurus, L. pictoense, L. plicatum, L. punctatum, L. rhodumnense, L. rimosum, L. scutatum, L. selaginoides, L. serpentigerum, L. sternbergii, L. tetragonum, L. undulatum, L. veltheimianum, L. veltheimii, L. volkmannianum, L. wortheni заставляет напрячься и постараться выявить основные признаки различий. Тем более, что часто новые виды возникали как сугубо субъективное мнение авторов. Описания некоторых из вышеприведенных видов можно посмотреть в работе Б. Томаса [2].
Стернберг в свое время внес немалую сумятицу в умы палеоботаников, объединив под именем L. dichotomum два разных вида лепидодендронов L. obovatum и L. longifolium, поскольку первый имеет межлистовые парихны, второй – нет. К несчастью, Стернберг в 1820 году под своим видом L. оbovatum объединял также такие очевидно различные виды, как L. аculeatum и L. mannebachense (1838) Пресля, что привело к возникновению более, чем вековой таксономической и номенклатурной путаницы в литературе. Němejc (1947) разрешил только таксономическую часть проблемы, когда подробно проанализировал различия между L. aculeatum и L. оbovatum и обеспечил их синонимичность. К сожалению, он сохранил сбивающее с толку имя L. оbovatum. Эта номенклатурная проблема сохранилась до 1970 года, когда Thomas предложил L. mannebachense Presl. как подходящее имя для замены L. оbovatum. Он же предложил установить синонимичность видов L. оbovatum и L. aculeatum [1].
Я же долго не мог понять различий между этими двумя видами, хаотически называя обоватумом то один образец, то другой. И даже теперь, посмотрев на иллюстрации относительно свежей публикации чешского палеоботаника Oplustil по онтогенезу (стадиям развития) Lepidodendron mannebachense (синоним L. obovatum), могу сказать, что внешне форма листовых подушек одного и того же вида имеет достаточно различную форму на разных стадиях развития растения.
Что еще можно сказать об особенностях лепидодендронов? Ствол имел довольно толстую в сравнении с современными деревьями кору, которая содержала в большей мере лигнин, чем целлюлозу. Огромные стволы поддерживали вертикальное положение с помощью стигмарий, корнеподобных структур, которые имели неглубокое заложение в грунте и боковые отростки, сбрасываемые со временем подобно листьям.
Существует ложный стереотип об исключительно дихотомическом ветвлении лепидодендрона. Напоминаю, что при дихотомическом ветвлении конус нарастания (апекс) делится надвое, вновь образующиеся апексы также делятся надвое и так далее. Различают так называемое изотомическое дихотомическое ветвление (образующиеся ветви равны по размеру) и анизотомическое (при котором образующиеся ветви не равны).
Ювенильная стадия развития растения завершалась первой дихотомией. Таким образом, до момента первого разделения апекса (дихотомии) растение выглядело этаким столбиком с мохнатой верхушкой (см. иллюстрацию).

Иллюстрация из работы [1]
В соответствии с современными представлениями, лепидодендрон имел боковые фертильные побеги, нарушающие строгую дихотомию. На образцах лепидодендронов часто наблюдаются псевдомонопоидальные ветви (у растений с моноподиальным типом строения побега верхушечная почка сохраняется на протяжении всей жизни побега, а на самом побеге многократно образуются боковые побеги более высокого порядка).
Большинство образцов, например, Lepidodendron mannebachense демонстрируют наличие так называемых основных ветвей, появляющихся вследствие изотомической или анизотомической дихотомии, которые создают основу кроны дерева. Основные ветви держат на себе более мелкие псевдомоноподиальные и строго анизотомические боковые ветви, плотно размещенные на расстоянии нескольких сантиметров друг от друга.

Иллюстрация из работы [1]
Эти боковые ветви в свою очередь могут поддерживать еще более малые следующего порядка, завершающиеся спороносной шишкой (стробилом). Сочетание этих двух механизмов ветвления предполагают, что апекс ювенильного ствола этого лепидодендрона начинает дихотомически делиться и это продолжается до тех пор, пока не будет сформирована вся крона и не прекратится рост дерева. Если Lepidodendron mannebachense полностью вырастал и только после этого появлялись спороносные шишки, то такой вид, как Diaphorodendron размножался непрерывно, достигнув определенного возраста (а не в самом конце жизни). Среднее количество дихотомий точно установить не удается, однако они повторяются в среднем через каждые полметра вдоль оси ветви.
Отношение высоты листовой подушки к ее ширине (как правило, больше единицы для лепидодендронов) тоже варьирует на одном и том же растении. Размеры подушек тем крупнее, чем старше кора (или порядок ветви). Соотношение высота/ширина уменьшается на более молодых ветвях и наблюдается прямая, хоть и не линейная, зависимость размеров подушки от диаметра стебля. То же можно сказать о размере листьев. Чем выше порядок ветвления, тем короче листья. Отсутствие листьев на большей части образцов ветвей за исключением временных боковых побегов говорит о том, что листья имели короткий функциональный период в жизни растения и довольно быстро сбрасывались. То есть, на конечной стадии жизненного цикла растения листья присутствовали только на боковых фертильных побегах. Эти листья также сбрасывались, очевидно, после того, как спороносные шишки созревали.Интересно, что для Lepidodendron mannebachense характерно наличие листьев на стебле или ветви только в узкой зоне в непосредственной близости от апекса.

В заключение хотелось бы заметить, что иногда находимые образцы не вписываются в схемы, приведенные в публикации. Например, ниже показан образец, у которого листовые рубцы как бы частично прикрыты элементами листовой подушки.
[несуществующая фотография]
А в этих случаях, листовой рубец закрыт своего рода щитком, прикрывающим и парихны, и проводящий пучок.(В первом случае видны следы вторичного роста - широкие расстояния между листовыми подушками).


Литература
1. OPLUŠTIL, S. 2010. Contribution to knowledge on ontogenetic developmental stages of Lepidodendron mannebachense Presl, 1838. Bulletin of Geosciences 85(2), 303–316 (7 figures). Czech Geological Survey, Prague. ISSN 1214-1119. June 30, 2010.
2. THOMAS, B.A. 1970. Epidermal studies in the interpretation of Lepidodendron species. Palaeontology 13(1), 145–173.
Ключевые слова (тэги): лепидодендрон, парихна, Lepidodendron, дихотомия
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 18 ноября 2011 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://ammonit.ru./text/290.htm#47277">http://ammonit.ru./text/290.htm#47277</a> |
|
|||||
Отличная публикация!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 19 ноября 2011 года в 14:05:02 |
|||||
 Добавить в избранное
Добавить в избранное 


|
<a href="http://ammonit.ru./text/290.htm#47282">http://ammonit.ru./text/290.htm#47282</a> |
|
|||||
Спасибо, Андрей, этот материал скорее дополняет предыдущий по лепидодендронам. | ||||||
|
комментарий 2 уровня |
Комментарий создан 19 ноября 2011 года в 15:00:38 |
|||||

|
<a href="http://ammonit.ru./text/290.htm#47290">http://ammonit.ru./text/290.htm#47290</a> |
|
|||||
С интересом все прочитал и спасибо за первый рисунок. | ||||||
|
комментарий 1 уровня |
Комментарий создан 19 ноября 2011 года в 17:26:10 |
|||||
|
<a href="http://ammonit.ru./text/290.htm#50833">http://ammonit.ru./text/290.htm#50833</a> |
|
|||||
Были и без лигульные формы | ||||||
|
комментарий 2 уровня |
Комментарий создан 25 декабря 2011 года в 14:42:57 |
|||||

|
<a href="http://ammonit.ru./text/290.htm#50856">http://ammonit.ru./text/290.htm#50856</a> |
|
|||||
Дмитрий, у меня к Вам просьба, дайте комментарий по роли и функциональным нагрузкам лигулы. Для чего она вообще нужна была растениям? | ||||||
|
комментарий 3 уровня |
Комментарий создан 25 декабря 2011 года в 18:07:49 |
|||||
|
<a href="http://ammonit.ru./text/290.htm#50863">http://ammonit.ru./text/290.htm#50863</a> |
|
|||||
Одна из версий что это орган для поглащения дополнительной влаги из атмосферы. | ||||||
|
комментарий 4 уровня |
Комментарий создан 25 декабря 2011 года в 18:58:03 |
|||||
|
<a href="http://ammonit.ru./text/290.htm#50866">http://ammonit.ru./text/290.htm#50866</a> |
|
|||||
Спасибо, я давно этим интересуюсь, но внятной версии не находил. | ||||||
|
комментарий 5 уровня |
Комментарий создан 25 декабря 2011 года в 19:23:13 |
|||||