|
|
 |
|
Аммониты и стратиграфия кимериджского и волжского ярусов (а также эволюция краспедитид)
Автор: MHorn
Рубрика: Палеонтологические публикации
По примеру коллег выкладываю здесь ссылку на свою часть, опубликованную в Трудах ГИН:
Рогов М. А. (2017) Аммониты и инфразональная стратиграфия кимериджского и волжского ярусов юга Московской синеклизы // Труды ГИН РАН. Вып. 615. С.7-160.
pdf: http://mmtk.ginras.ru/pdf/rogov2017_km_vlg_moscow.pdf
Однако размещать тут целиком всю работу на полторы сотни страниц в виде публикации – это явный перебор, так что ограничусь лишь одним разделом, посвящённым эволюции поздневолжских краспедитид. Остальное же – описание разрезов, зон, подзон и биогоризонтов, а также аммонитов (и 20 фототаблиц) можно найти в пдфе по ссылке выше.
Краткий обзор особенностей эволюции эудемичных поздневолжских краспедитид в Среднерусском море
Поздневолжские краспедитиды, обитавшие в Среднерусском море, представляют собой благодатный материал для изучения особенностей эволюции аммонитов: два подсемейства, к которым относится подавляющее большинство всех находок поздневолжских форм (Garniericeratinae и Craspeditinae), представлены быстро эволюционирующими эудемичными филолиниями, тогда как роль таксонов-иммигрантов на протяжении практически всего поздневолжского времени оставалась невелика. К настоящему времени собран представительный материал по обоим подсемействам из разнофациальных отложений, накапливавшихся на разных глубинах при различном удалении от берега.
Первые гарниерицератины (Kachpurites) появились в конце фазы Nikitini (гемера nikitini), сразу после кризиса, который в бореальных бассейнах привёл в к исчезновению сообществ с доминированием мегаконхов дорзопланитид. Эти древнейшие Kachpurites, представленные почти исключительно микроконхами диаметром 2-3 см (Рогов, 2015, рис. 1, A-B), характеризовались очень низкой изменчивостью. Раковины этих аммонитов, как правило, встречаются в огромных количествах, образуя ракушняки во всех районах распространения вида (Ярославская обл., Ульяновская обл., Татарстан, Москва и Подмосковье). Средневолжские кашпуритесы характеризуются эволютным навиванием раковины, простой скульптурой и лопастной линией – признаками, которые часто сопутствуют резким эволюционным перестройкам у аммонитов (Guex, 2001, 2006). Древнейшие кашпуритесы необычайно сходны по форме раковины и характеру скульптуры с внутренними оборотами Laugeites и, судя по всему, произошли от них путём педоморфоза. К началу поздневолжского времени облик гарниерицератин не претерпел заметных изменений – K. evolutus отличались от более древних кашпуритесов характером скульптуры, но имели сходные размеры и форму раковины.
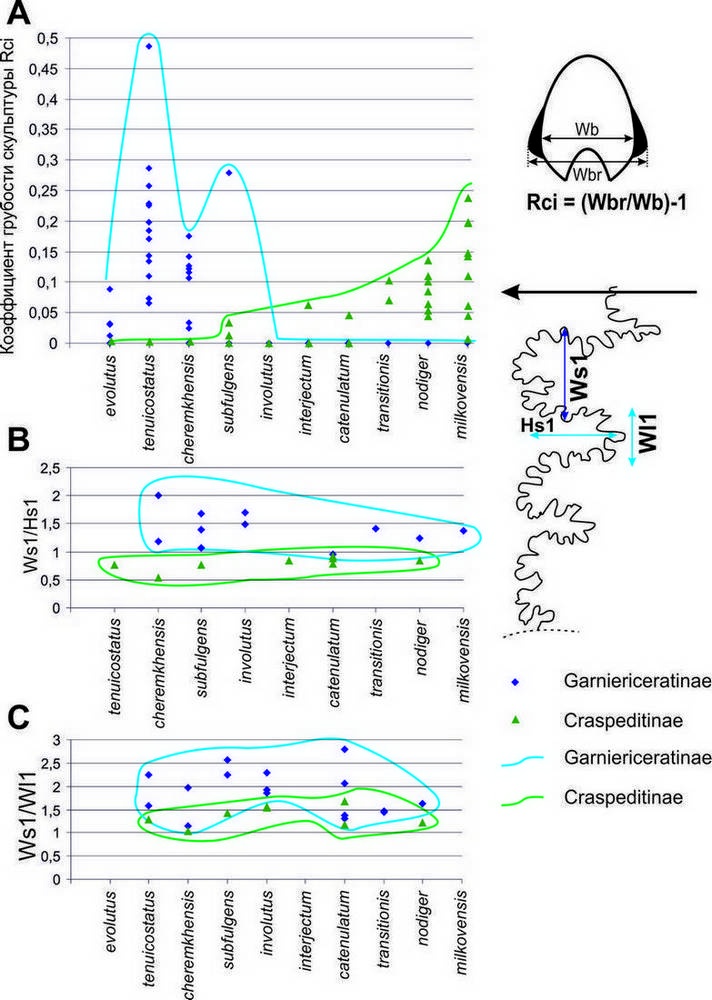
Рис. 1. Изменение некоторых параметров скульптуры (A - грубость первичных рёбер / бугорков) и лопастной линии (B, C - относительные размеры лопасти L к соседнему седлу) в эволюции поздневолжских гарниерицератин и краспедитин.
У более поздних K. tenuicostatus, характеризовавшихся наиболее грубой скульптурой (рис. 1) и наибольшей изменчивостью среди волжских краспедитид, регулярно начинают встречаться крупные (10-15 см и более) морфы, которые известны как в Среднем Поволжье (Кашпир, Марьевка), так и в Москве и Подмосковье (где они, как правило, представлены фрагментами). Такие крупные экземпляры регулярно встречаются и у более молодых видов кашпуритесов, но их находки приурочены преимущественно к Ярославскому Поволжью, тогда как в других регионах они редки или отсутствуют. Начиная с K. cheremkhensis изменчивость скульптуры кашпуритесов начинает сокращаться, минимальна она у самого позднего вида данного рода K. involutus. В фазу Fulgens гарниерицератины резко преобладали во всех изученных комплексах, составляя, как правило, более 90% находок (рис. 2, см. также Рогов и др., 2015).
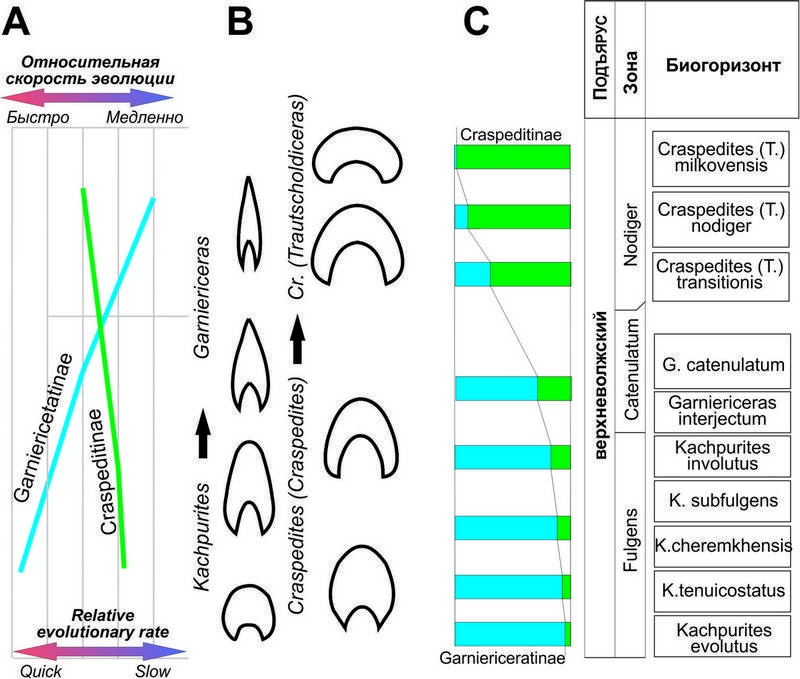
Рис. 2. Изменение относительной скорости эволюции гарниерицератин и краспедитин (А), схема изменения поперечного сечения раковины гарниерицератин и краспедитин (В) и усреднённые значения относительного обилия гарниерицератин (показаны синим) и краспедитин (показаны зелёным) в поздневолжское время (С).
На фоне высокого обилия гарницерицератины фазы Fulgens быстро эволюционировали, пройдя путь от сильно скульптированых платиконов до практически лишённых скульптуры дискоконов. В начале фазы Catenulatum от Kachpurites происходят первые Garniericeras, обладающие практически идентичной с поздними кашпуритесами скульптурой и формой раковины (не случайно по форме раковины самый молодой вид кашпуритесов K. involutus уже попадает в основном в поле, занятное родом Garniericeras –рис. 3).
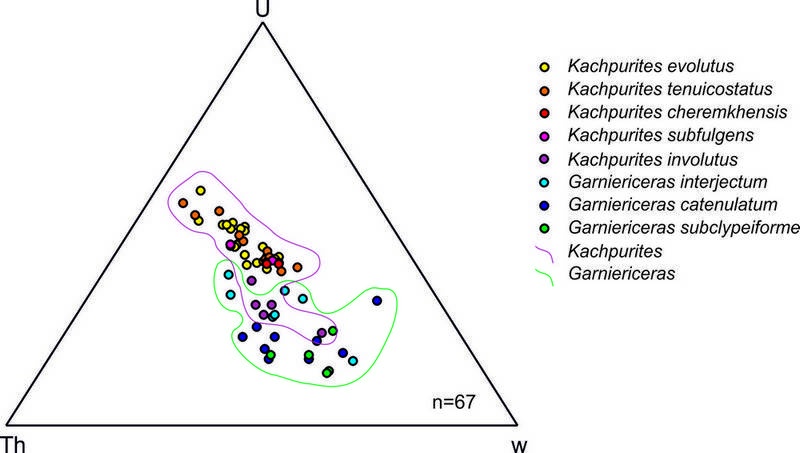
Рис. 3. Изменение параметров формы раковины по методу морфообласти Вестерманна (Ritterbush, Bottjer, 2012) в эволюции поздневолжских гарниерицератин. Треугольная диаграмма составлена с помощью программы PAST. Конечные морфообласти углов диаграммы: Th – серпентиконы, U – сфероконы, W – оксиконы.
Поздние кашпуритесы от ранних гарниерицерасов отличаются одной единственной особенности – присутствуием у гарниерицерасов киля, сначала на внутренних оборотах (G. interjectum), а затем и на КЖК (G. catenulatum; у этого вида в качестве очень редкого варианта изменчивости отмечаются находки раковин без киля, практически не отличимые от кашпуритесов). Скульптура у гарниерицерасов почти отсутствует – лишь у G. interjectum наблюдаются тонкие нитевидные рёбрышки в верхней части оборота, аналогичные рёбрам поздних кашпуритесов, и иногда на боковой стороне раковины могут присутствовать слабо выраженные более грубые рёбра (только у G. catenulatum). Интересно отметить, что если у G. interjectum преобладали сравнительно небольшие по размеру раковины, то у их потомков G. catenulatum и средний и максимальный размеры раковин существенно возрастают. У самых поздних представителей рода Garniericeras (и подсемейства гарницерицератин) G. subclypeiforme раковина становится очень узкой, оксиконической, с очень узким умбиликусом. Но если ранние гарниерицерасы всё ещё встречались достаточно часто, то G. subclypeiforme в большинстве комплексов сравнительно редки (исключение здесь составляет разве что комплекс из биогоризонта transitionis Мильково) и становились всё более редкими к концу фазы Nodiger, где их находки составляют первые проценты (рис. 2 С). Последние гарниерицерасы известны из верхней зоны волжского яруса Singularis, где в биогоризонте pulcher отмечаются единичные находки Garniericeras плохой сохранности, идентифицируемые только по особенностям строения лопастной линии. Гарниерицератины окончательно исчезли незадолго до конца поздневолжского времени, не оставив потомков.
Таксономическое положение микроконхов из группы “Craspedites” ivanovi Geras., спорно. Эти аммониты в массовом количестве появляются вблизи основания подзоны Rosanovi зоны Virgatus и изредка встречаются в зоне Nikitini. По форме раковины и характеру скульптуры они близки микроконхам поздневолжских Craspedites, но отличаются более узким умбиликусом. По всей видимости, эта своеобразная группа микроморфных дорзопланитид, произошедшая путём педоморфоза от Serbarinovella (Митта, 1993), не связана с настоящими краспедитидами (Rogov, Zakharov, 2009). Первые несомненные краспедитины (Craspedites (C.) praeokensis и сопутствующие этому виду микроконхи) появляются в начале поздневолжского времени. Макроконхи ранних краспедитин близки по форме раковины, размеру и скульптуре поздним Laugeites из подзоны Nikitini, отличаясь от них лишь особенностями скульптуры внутренних оборотов, для которых характерны короткие рёбра в верхней части боковой стороны, и более «краспедитовым» обликом лопастной линии, хотя подобная линия иногда присутствует и у лаугеитин (у всех Epilaugeites и некоторых Laugeites). Таким образом, можно предполагать происхождение Craspedites (Craspedites) от Laugeites. Появление краспедитид сопровождалось уменьшением средних и максимальных размеров раковин по сравнению с лаугетеинами (этот процесс начался ещё в середине фазы Nikitini, когда в высоких широтах Северного полушария произошло повсеместное уменьшение размеров раковины у дорзопланитид). В дальнейшем в течение поздневолжского времени размер раковины краспедитин постоянно уменьшался (рис. 4); наиболее чётко эта тенденция проявляется у представителей подсемейства, населявших Среднерусское море, но прослеживается также и у арктических представителей рода (в линии C. (Craspedites) – C. (Taimyroceras)).
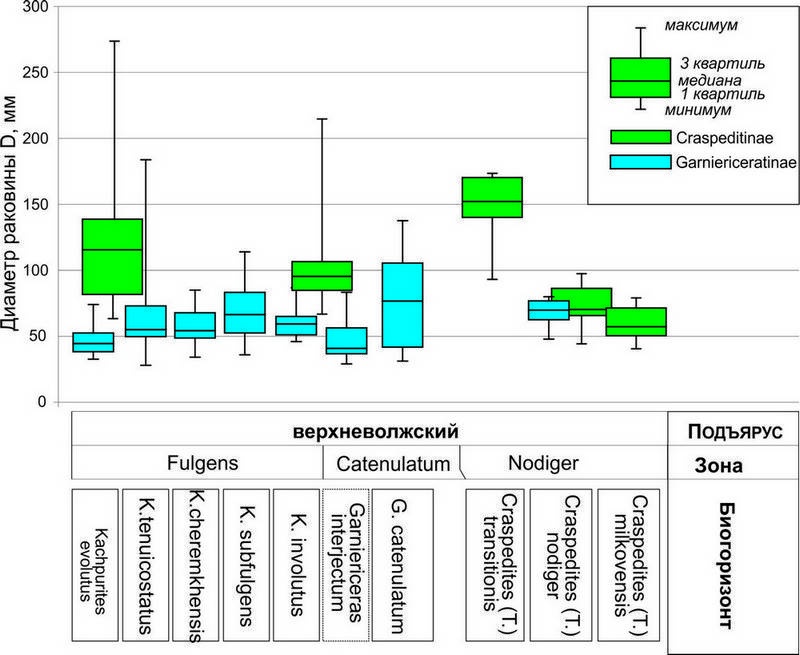
Рис. 4. Диаграммы с усами (чёрная горизонтальная линия – медиана, блоки вокруг неё – 1 и 3 квартили), показывающая изменение размеров раковины в эволюции эудемичных филолиний макроконхов гарниерицератин и краспедитин Среднерусского моря.
Скорость эволюции и изменчивость у макроконхов ранних краспедитин (подрод Craspedites s. str.) были невелики (рис. 2 20А). За исключением довольно сильного варьирования конечных размеров раковины остальные параметры формы раковины и скульптуры у этих аммонитов были практически идентичны; в качестве одного из вариантов изменчивости скульптуры можно отметить лишь сравнительно нечастое появление первичных рёбер на жилых камерах. Ситуация резко изменилась в конце поздневолжского времени, в фазу Nodiger, когда появился эудемичный подрод С. (Trautscholdiceras) В это время в большинстве комплексов краспедитины стали заметно преобладать над гарниерицератинами, у них увеличилась изменчивость, грубость скульптуры и скорость появления новых таксонов (рис. 1-2). Однако, в конце фазы Nodiger С. (Trautscholdiceras) полностью вымерли, и в вышележащей зоне Singularis краспедитин нет, а в интервале между зонами Nodiger и Singularis в бассейне р. Черёмухи встречен единственный экземпляр краспедитид, близкий к Craspedites (Taimyroceras).
Как видно из приведённого обзора, несмотря на то, что обе эудемичные филолинии поздневолжских краспедитид Среднерусского моря населяли один и тот же бассейн, эволюционные тенденции в этих группах были резко различны практически по всем характеристикам (относительное обилие, скорость эволюции, диапазон изменчивости, эпизоды появления грубо скульптированных форм). Интересно, что некоторые из отмеченных для аммонитов Среднерусского моря тенденций проявились и в эволюции краспедитид, населявших другие бореальные бассейны. Так, очень близко направление эволюции гарниерицератин и субкраспедитин (Рогов, 2014 a), обитавших в поздневолжское время преимущественно в северо-западной Европе. Изменения формы раковины и скульптуры в линии Craspedites sp. str. – C. (Trautscholdiceras) близки к изменениям в линии Craspedites s. str. – C. (Taimyroceras), хотя усиление первичных рёбер у таймыроцерасов было выражено в несколько более слабой степени, чем у траутшольдицерасов. С учётом того, что перечисленные выше группы аммонитов эволюционировали не только в разных бассейнах, но и в бассейнах, имеющие противоположные тенденции развития в терминальной юре (моря северо-западной Европы и европейской России регрессировали, а в большей части Арктики, наоборот, происходило постепенное повышение уровня моря) выявление факторов, влияющих на такие особенности эволюции краспедитид, представляет собой непростую задачу. По всей видимости, основную роль тут играли биотические факторы среды.
Ключевые слова (тэги): аммониты, эволюция, стратиграфия
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 28 сентября 2017 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://ammonit.ru./text/2081.htm#307422">http://ammonit.ru./text/2081.htm#307422</a> |
|
|||||
Я бы добавил, что в этой замечательной публикации приведены описания множества хорошо знакомых московским любителям верхнеюрских разрезов, таких как Еганово, Мневники, Фили (Кунцево), Рыбаки и т.д. Я думаю, что это будет интересно читателям Аммонит.ру | ||||||
|
комментарий 1 уровня |
Комментарий создан 28 сентября 2017 года в 22:11:28 |
|||||
 Добавить в избранное
Добавить в избранное 


|
<a href="http://ammonit.ru./text/2081.htm#307425">http://ammonit.ru./text/2081.htm#307425</a> |
|
|||||
Интересно, и очень объемно, новые таксоны для средней волги даже) Dorsoplanites сf. multiconstrictus у себя в сборах уже нашел, есть такие часторебристые тонкие по вентру, надо поискать Zaraiskites cf. kuteki :) | ||||||
|
комментарий 1 уровня |
Комментарий создан 28 сентября 2017 года в 23:36:54 |
|||||

|
<a href="http://ammonit.ru./text/2081.htm#307459">http://ammonit.ru./text/2081.htm#307459</a> |
|
|||||
Очень интересно.
| ||||||
|
комментарий 1 уровня |
Комментарий создан 29 сентября 2017 года в 15:35:45 |
|||||
|
<a href="http://ammonit.ru./text/2081.htm#307492">http://ammonit.ru./text/2081.htm#307492</a> |
|
|||||
Я там не бы с начала 90х, да и там в основном наименее интересный интервал, в котором никакой стратиграфии не сделать - зона Panderi | ||||||
|
комментарий 2 уровня |
Комментарий создан 29 сентября 2017 года в 22:59:24 |
|||||

|
<a href="http://ammonit.ru./text/2081.htm#307697">http://ammonit.ru./text/2081.htm#307697</a> |
|
|||||
Зона Panderi там на 6-ти метровой высоте, а в уровне летней воды непонятно кимеридж или оксфорд. В интервале 2-х метров (1м над летним уровнем воды и 1м под ним)встречается много Amoeboceras-ов в виде отпечатков в глине, иногда попадаются Ringsteadia в слоях доступных зимой. | ||||||
|
комментарий 3 уровня |
Комментарий создан 2 октября 2017 года в 10:07:47 |
|||||
|
<a href="http://ammonit.ru./text/2081.htm#307712">http://ammonit.ru./text/2081.htm#307712</a> |
|
|||||
надо бы туда выбраться - как раз сейчас нужно срочно отобрать образцы из пограничного интервала оксфорда и кимериджа | ||||||
|
комментарий 4 уровня |
Комментарий создан 2 октября 2017 года в 14:51:04 |
|||||