|
|
 |
|
Прижизненные повреждения на раковинах юрских аммонитов Центральной России
Автор: Alex
Рубрика: Палеонтологические публикации
Уважаемые читатели Аммонит.ру! В начале сентября вышла из печати книга Труды Геологического института РАН. Выпуск № 615: Юрские отложения юга Московской синеклизы и их фауна, подготовленная коллективом авторов из ГИНа, ПИНа и МГУ.
В сборнике вышли две моих статьи - одну из них я разместил несколько дней назад: Анатомия и некоторые аспекты палеобиологии аммоноидей: современные данные.
Ниже представлен текст второй статьи, она называется Прижизненные повреждения на раковинах юрских аммонитов Центральной России, в виде pdf-файла ее можно скачать на ResearchGate: Sublethal injuries on the shells of Jurassic ammonites from Centra Russia. В ней дается короткий обзор прижизненных повреждений и аномалий развития раковин аммонитов и приводятся данные по распространенности повреждений и аномалий в келловейских и оксфордских отложениях трех местонахождений (Михайлов, Дубки и Марково).
----------------------------------------------
Прижизненные повреждения на раковинах юрских аммонитов Центральной России
Мироненко А.А., Геологический институт РАН, Москва
Прижизненные повреждения и патологии развития на раковинах аммонитов хранят информацию об образе жизни этих животных и о пищевых цепях, в которые аммониты были включены. Данные о том, какие травмы получали аммониты, с какими хищниками или паразитами они сталкивались, позволяют реконструировать не только их образ жизни, но и лучше понять как были устроены экосистемы, в которых они обитали. Однако, для этого нужно располагать не только информацией о том, какие типы повреждений и палеопатологий можно найти на аммонитах в целом, но и на каких именно аммонитах и как часто встречаются те или иные повреждения.
В данной публикации описываются прижизненные повреждения и аномалии, обнаруженные на раковинах аммонитов из четырех различных стратиграфических горизонтов келловея и оксфорда трех местонахождений Центральной России (Михайлов, Дубки и Марково). Среди изученных палеопатологий большая часть связана с повреждениями, вызванными атаками хищников, однако встречаются и искривления раковин в результате поселения обрастателей. Причины возникновения некоторых патологий остаются неясными (это могут быть инфекции, паразиты или неблагоприятные условия среды).
Наибольшее число прижизненных повреждений, связанных с атаками хищников, обнаружено на раковинах Binatisphinctes и Kosmoceras из среднего и верхнего келловея Михайлова. Доля поврежденных раковин среди этих аммонитов составляет около 16%. Также очень велико число патологий у верхнекелловейских Quenstedtoceras из Дубков (около 14%), но среди них встречаются не только травмы, но и аномалии, вызванные прикреплением раковин двустворок. В оксфорде поврежденные раковины встречаются реже. В среднем, в келловее Михайлова и Дубков около 10% раковин имеют те или иные патологии, в оксфорде Михайлова и Марково поврежденных или аномальных раковин в среднем около 5%. Снижение числа травм и патологий в оксфорде связано, по-видимому, с обеднением экосистем по сравнению с келловеем (число родов аммонитов, обитавших в Среднерусском море в это время также резко сократилось).
Установлено, что раковины разных аммонитов подвергались действию различных факторов, вызывавших патологии. Так, на раковинах Erymnoceras и Funiferites встречаются укусы, интерпретированные как следы атак рыб-дурофагов, похожие, но не идентичные повреждения встречаются и на оксфордских Amoeboceras. На раковинах Binatisphinctes и Kosmoceras встречаются в основном следы небольших укусов, но вызывавших серьезные травмы мантийного края (диагностирующиеся по шрамам на построенной после повреждения части раковины). Эти различия свидетельствуют о том, что разные роды аммонитов занимали разные экологические ниши (вероятно, связанные с разной глубиной обитания) и сталкивались с разными хищниками и условиями среды.
Ключевые слова: прижизненные повреждения, палеобиология, аммониты, аномалии развития, паразиты.
Введение
Раковины аммонитов несут на себе следы различных событий, происходивших с их обладателями, в том числе и таких, как нападения хищников, травмы, болезни и воздействие неблагоприятных условий окружающей среды. В результате нападений и травм на раковинах оставались характерные повреждения, а болезни или неблагоприятные условия приводили к формированию различных аномалий в ходе строительства раковины. Изучение этих палеопатологий помогает понять палеобиологию и палеоэкологию аммонитов: уточнить их анатомию, лучше понять механизм строительства раковины и выяснить, в какие пищевые цепи были включены эти моллюски.
К сожалению, интерпретация палеопатологий аммонитов и поиск вызвавших их причин являются довольно сложными задачами. Следы прижизненных травм и болезней легко отличить от посмертных или смертельных повреждений благодаря тому, что первые имеют характерные признаки залечивания повреждённой области или продолжения роста раковины. Однако, часто бывает сложно установить причину возникновения того или иного повреждения, вплоть до того, что в некоторых случаях остается неясным, связана ли патология с каким-либо внешним воздействием или с процессами, протекавшими в организме самого моллюска. В связи с этим, различные повреждения и аномалии на раковинах обычно классифицируют по внешним признакам, оставляя за скобками формальной классификации возможные причины их появления. Самой распространенной является классификация по так называемым типам «forma aegra» (Hölder, 1956; Hengsbach, 1996; Keupp, 2012; Hoffmann, Keupp, 2015), что можно перевести как «болезненная форма», примеры таких типов будут приведены ниже.
Формальная классификация удобна для описания раковин с видимыми патологиями, но при поиске причин возникновения этих патологий нужно помнить, что одно и то же повреждение (например, скол устьевого края) могло быть вызвано в разных случаях совершенно разными причинами, в то время как укус одного и того же хищника в разных ситуациях мог оставлять на раковинах совершенно разные следы — в зависимости от угла и области атаки и от того, был при этом поврежден край мантии аммонита или нет. Дело в том, что наиболее заметные палеопатологии, такие как длинные шрамы или утерянные скульптурные элементы (исчезнувшие ряды шипов или киль), приводящие к существенному изменению внешнего вида раковин, возникали из-за того, что при повреждении мантийного края, ответственного за формирование новых участков раковины, на время или навсегда нарушалось его функционирование. У аммонитов каждый участок мантийного края отвечал за строительства определенного участка раковины, и если участок, ответственный, к примеру, за формирование шипов оказывался утерянным, то в дальнейшем этот ряд шипов уже не формировался. Таким образом, в случае, если один и тот же хищник успевал ухватить аммонита за край мантии или же только за край раковины, эффект оказывался принципиально разным: в первом случае форма раковины, построенной после этого инцидента могла оказаться существенно искажённой, во втором случае все ограничивалось небольшим повреждением только в месте укуса.
Кроме того, некоторые варианты формальных типов повреждений связаны с определёнными элементами скульптуры, к примеру, forma aegra undaticarinata и forma aegra chaotica связаны с аномальным развитием киля на раковинах, соответственно, они могут наблюдаться только у аммонитов, имеющих выраженный киль. На раковинах другой морфологии, не имеющих киля, эти формальные типы не могли возникнуть даже при воздействии тех же самых факторов.
Разные таксоны аммонитов демонстрируют различную частоту палеопатологий, более того, процент поврежденных или аномальных раковин может различаться в несколько раз даже в относительно одновозрастных популяциях одного и того же таксона (Keupp, 2012). Сравнение частоты и типов повреждений, наблюдающихся на раковинах различных аммонитов в одном и том же местонахождении помогает уточнить образ жизни и области обитания этих животных. К примеру, высокий процент повреждений на раковинах определенного вида аммонитов, вызванных атаками ракообразных, может свидетельствовать о демерсальном образе жизни представителей этого таксона. Также важно сравнение прижизненных повреждений различной степени тяжести (то есть затрагивающих и не затрагивающих мантийный край). Несомненно, аммониты легче переживали травмы, не затрагивавшие их мягкое тело, поэтому высокий процент повреждений мантии может свидетельствовать о том, что в данном случае аммониты столкнулись с хищником, к противостоянию с которым они не были достаточно хорошо адаптированы.
К сожалению, сравнение палеопатологий раковин у разных таксонов аммонитов имеет свои ограничения. Сравнивая два таксона, у одного из которых залеченные повреждения часты, а у другого редки, мы не можем однозначно судить о причинах такого распределения. Возможно, что представители одного таксона чаще подвергались атакам хищника, но также не исключено, что в силу каких-либо причин у аммонитов, имеющих меньшее количество залеченных травм была значительно выше летальность в результате нападений. Для того, чтобы сравнить эти версии, необходимо параллельное изучение раковин с летальными, незалеченными повреждениями. Но во многих местонахождениях сделать это крайне трудно по нескольким причинам: во-первых, довольно сложно отличить смертельное повреждение, нанесенное хищником, от посмертного разрушения раковины падальщиками или абиотическими факторами, во-вторых, во многих местонахождениях плохо сохраняются жилые камеры, на которых как раз и можно найти следы летальных повреждений.
Тем не менее, изучение палеопатологий и сравнение их распространения между таксонами позволяет многое понять в палеоэкологии и палеобиологии аммонитов. В данной публикации представлены новые данные по прижизненным повреждениям и аномалиям развития, обнаруженным на раковинах юрских (келловейских и оксфордских) аммонитов Центральной России. Аммониты происходят из следующих местонахождений: Марково (Раменский район Московской области), Михайлов (карьеры в окрестностях одноимённого города Михайловского района Рязанской области) и Дубки (заброшенный карьер близ пгт. Дубки Саратовского района Саратовской области). Детальное описание разрезов, расположенных в окрестностях г. Михайлов, приведено в настоящем сборнике (Тесакова и др., 2017), карьер Дубки описан в ряде публикаций, наиболее полные данные приведены в статье Д.Н. Киселёва с соавторами (Kiselev et al., 2013). Разрез, расположенный на берегу р. Москвы в д. Марково хорошо известен специалистам (см. Mironenko, 2015, Fig.1), но до настоящего времени его описание не опубликовано, что в значительной степени связано с характером обнажения: верхнеоксфордские слои здесь обнажаются только зимой при низком уровне воды в р. Москвы, и обычно вскрываются ямами. Данные по прижизненным повреждениям у келловейских аммонитов из Михайлова и Дубков ранее приводились в литературе (Keupp, 2012 и Сельцер, 2001; Larson, 2007 соответственно), но палеопатологии оксфордских аммонитов из Михайлова и Марково до сих пор не исследовались.
Интерпретация прижизненных повреждений раковин аммонитов.
Прижизненные повреждения и аномалии развития раковины аммонитов могли быть вызваны самыми разными причинами — от атак хищников до генетических, инфекционных или паразитарных заболеваний.
Самыми распространёнными прижизненными повреждениями являются следы нападений различных хищников (см. Keupp, 2012 и Hoffmann, Keupp, 2015). Изучение таких повреждений всегда вызывало большой интерес, тем более, что для определения роли аммонитов в трофических цепях важно понимать, кто именно на них охотился. Залеченные прижизненные повреждения помогают в разрешении этого вопроса, хотя их интерпретация зачастую является сложным делом. В палеонтологической литературе в качестве потенциальных агрессоров, нападавших на аммонитов, называют практически все группы хищных позвоночных и беспозвоночных, обитавших в палеозойских и мезозойских морях (Klompmaker et al. 2009; Keupp, 2012; Hoffmann, Keupp, 2015). Среди позвоночных это различные рептилии и рыбы, среди беспозвоночных — ракообразные, колеоидеи, наутилиды и сами аммоноидеи. К сожалению, во многих случаях практически невозможно определить, кто именно из многочисленных и разнообразных хищников ответственен за данное повреждение раковины аммонита. Однако, в отношении некоторых типов повреждений существуют вполне аргументированные предположения.

Рис. 1. Залеченные прижизненные повреждения устьевого края (forma aegra substructa), возникшие, скорее всего, при нападении рыб. A. Прижизненное повреждение на фрагменте раковины Amoeboceras alternoides, верхний оксфорд, Марково, Московская обл. Черная стрелка отмечает повреждение, белая — участок морщинистого слоя (затылочная прикрепительная область, см. Mironenko, 2015). Масштабный отрезок 5 мм. B – Залеченное повреждение (отмечено стрелками) на раковине Funiferites allae, верхний келловей, Михайлов, Рязанская обл. B1 – вид сбоку, B2 – вид с вентра. Масштабные отрезки равны 1см. C - Залеченное повреждение (отмечено стрелкой) на раковине Erymnoceras doliforme, средний келловей, Михайлов, Рязанская область. Масштабный отрезок 5 мм. D - Залеченное повреждение на раковине современного Nautilus pompilius. Масштабный отрезок 1 см.
Множественные выбоины по краю временного устья раковин (Рис.1) интерпретируются как следы атак костистых рыб (Hoffmann, Keupp, 2015). Точно такие же следы часто встречаются на раковинах современных наутилусов (Рис.1D) и исследователи неоднократно наблюдали момент их образования при нападении рыбы на устьевой край раковины (Saunders et al. 2010). В некоторых случаях эти выбоины имеют характерную полукруглую форму, в таком случае виновными в их появлении считают рыб из семейства Semionotidae, имевших многочисленные округлые зубы и явно бывших хищниками-дурофагами, которые вполне могли охотиться на аммонитов (Keupp, 2012; Hoffmann, Keupp, 2015).
Длинные узкие вырезы, начинающиеся от устья и идущие иногда до половины длины жилой камеры интерпретируются как следы атаки ракообразных, так как ни один другой хищник не имеет настолько длинного и мощного, и при этом узкого «инструмента», пригодного не для разламывания на куски, а для прорезания раковины (Kröger, 2002; Wani et al. 2012). Также следами атак ракообразных являются, вероятнее всего, симметричные прорезы обеих латеральных стенок по бокам устья (Рис.2).

Рис. 2. Залеченные прижизненные повреждения дорсо-латеральных участков раковин аммонитов. Повреждения узкие и почти симметричны, при этом вентральная сторона остается неповрежденной. Скорее всего, эти травмы были нанесены клешнями ракообразных. A. — почти симметричные дорсо-латеральные повреждения на раковине Cardioceras sp. (нижний оксфорд, Михайлов, Рязанская обл.). Масштабные отрезки равны 1 см. B. — дорсо-латеральные повреждения в пупковой части раковины Erymnoceras coronatum (средний келловей, Михайлов, Рязанская обл.). Масштабные отрезки равны 5 мм.
Сложнее определить подозреваемых в нанесении узких v-образных или точечных повреждений устьевого края: такой след может оставить и клюв наутилуса или кальмара, и клешня рака, а на довольно крупной раковине — и челюсти небольшой рыбы. Стоит отметить, что довольно часто такие небольшие повреждения сопровождаются на последующей части раковины хорошо заметным шрамом, что свидетельствует о травмировании мантийного края, формировавшего раковину (Рис.3A,B,D).

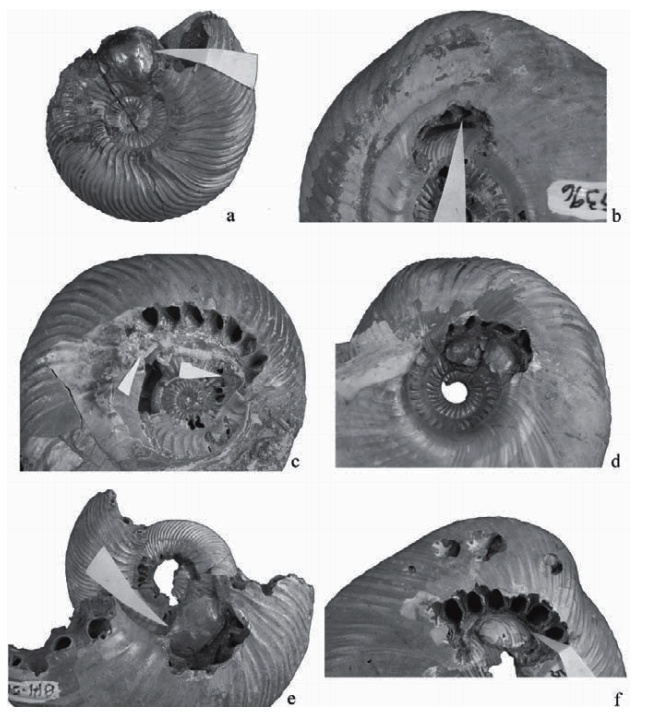
Рис. 3. Различные варианты аномального роста раковин вызванные травматическим повреждением мантии. A-C, E – Kosmoceras, средний келловей, Михайлов, Рязанская обл. D – Cardioceras, нижний оксфорд, Михайлов, Рязанская обл. A,B,D - forma aegra verticara, длинный шрам, присутствующий на построенной после травмы части раковины. Обратите внимание на разницу между травмированной (D1) и неповрежденной (D2) сторонами раковины: они выглядят как разные аммониты. C – forma aegra calcar, киль образующийся на раковине Kosmoceras из вентрального ряда шипов из-за утери второго ряда в результате травмы. E – раковина Kosmoceras, на которой из-за травмы исчезли оба вентральных ряда шипов и появился шрам по центру вентральной стороны. Длина масштабных отрезков 1 см.
Также в нанесении точечных или v-образных укусов могли быть виновны аммоноидеи с анаптиховым или ринхаптиховым типами челюстей (e.g. Lytoceratina, Phylloceratina). Вполне вероятно, что эти аммоноидеи могли охотиться на других аммонитов и своих сородичей, оставляя заметные повреждения на их раковинах. Но в юре Центральной России этих аммоноидей не было, здесь обитали только аммониты с аптиховым типом челюстного аппарата, мало пригодным к нанесению сильных укусов в край раковины. Некоторые исследователи полагают, что аммониты с аптиховым типом челюсти (Aptychophora согласно Engeser, Keupp, 2002) были исключительно микрофагами и планктонофагами (Kruta et al. 2011), однако, до тех пор, пока не будет детально изучено строение их верхней челюсти, эти выводы выглядят преждевременными. Тем не менее, укусить другого аммонита именно за край раковины, используя нижнюю челюсть аптихового типа, действительно было бы практически невозможно и из списков «подозреваемых» в нанесении большинства прижизненных повреждений юрских аммонитов Центральной России нужно исключить.

Рис. 4. Смертельные вентральные укусы на раковинах диморфной пары аммонитов подсемейства Hecticiceratinae, средний келловей, Михайлов, Рязанская обл. A – Sublunuloceras sp. [M], масштабный отрезок 1 см, B – Brightia sp. [m], масштабный отрезок 5 мм. В правой нижней части снимка веден устьевой вырост (ушко), устьевая часть жилой камеры разрушена при извлечении образца из породы.
Но один тип прижизненных повреждений может быть связан с атаками аммонитов с аптиховым типом челюсти — это вентральные укусы (Klompmaker et al. 2009; Andrew et al. 2010) (Рис.4). Хотя обычно в нанесении вентральных укусов подозревают колеоидей, нельзя исключать того, что их могли наносить самые разные хищники, приспособленные к охоте на аммонитов, так как эти укусы приходились в область прикрепления мускулатуры к раковине аммонита и в большинстве случаев оказывались смертельными. Аммониты для нанесения таких укусов вполне могли использовать аптихи в качестве своеобразной наковальни, нанося мощный удар по раковине жертвы верхней челюстью. Подобное предположение выглядит тем более вероятным, что челюстной аппарат более древних, триасовых аммоноидей был, по всей видимости, приспособлен именно к пробиванию, а не разрезанию добычи (Dagis, Weitchat, 1988). Залеченный вариант вентрального укуса называется forma aegra fenestra (от лат. «fenestra» - «окно») и довольно редко встречается на раковинах.
Однако, пример современных наутилусов показывает, что не всегда укусы в области устья бывают нанесены хищниками. Наутилусы иногда получают травмы во время охоты - от сопротивляющейся жертвы, или во время брачных турниров, когда самцы дерутся за самку (Несис, 2005). Но аммониты - не наутилусы, их половой диморфизм и их челюстной аппарат существенно отличались от таковых у современных наутилид. Как уже было сказано выше, челюстной аппарат аптихового типа не был пригоден для укусов в устьевой край раковины. Кроме того, самцы аммонитов за самку драться вряд ли могли и вообще вряд ли были драчливы: среди современных головоногих агрессивны самцы только тех видов, у которых размер самцов хоть ненамного, но превышает размер самок (Несис, 2005), а у аммонитов ситуация была обратной: самки были существенно крупнее самцов (см. Makowski, 1962), также как у современных осьминогов надсемейства Argonautoidea, у которых самцы за самок никогда не дерутся (Несис, 2005; Norman et al. 2002).
А вот травмы от защищающейся добычи аммониты вполне могли получать. Наутилусы иногда получают серьезные повреждения устьевого края, атакуя крабов и других ракообразных, которые защищаются своими сильными клешнями (Ward, 1981). Поэтому следы укусов ракообразных (а возможно, что в некоторых случаях и рыб) не всегда могут однозначно свидетельствовать о том, что хищник нападал на аммонита. Возможно, что в этой ситуации аммонит сам был агрессором, получившим травму от защищавшейся добычи.
Конечно, далеко не все палеопатологии раковин аммонитов связаны с укусами или травмами. Во многих случаях аномалии развития раковины были, по-видимому, вызваны болезнями, в том числе паразитарными, или неблагоприятными условиями окружающей среды. В литературе описаны своеобразные полукруглые выступы на внутренней поверхности раковин аммонитов, с помощью которых моллюски пытались изолировать каких-то некрупных паразитов (вероятно, червей), обитавших между раковиной и мантией (De Baets et al. 2011, 2015). Подобные «полужемчужины» не видны снаружи раковины, они обнаруживаются либо на срезах стенки раковины, либо их отпечатки в виде характерных углублений сохраняются на ядре. В юрских отложениях (в отличие от палеозойских) такие следы паразитов редки во всем мире, в Подмосковье один отпечаток подобной полусферы обнаружен на ядре жилой камеры поздневолжского Kachpurites (Mironenko, 2016, фиг.8E).
Также с инфекциями или деятельностью паразитов часто связывают образование различных вздутий на раковинах аммонитов в тех случаях, когда этим патологиям не предшествует явное повреждение раковины. Такие вздутия довольно часто встречаются на верхнекелловейских аммонитах Quenstedtoceras из местонахождения Дубки в Саратовской области (Larson, 2007). Также стоит упомянуть очень необычных эктопаразитов, обитавших на раковинах поздневолжских Kachpurites в Московском регионе — их деятельность приводила к существенному искривлению растущей части раковины и образованию вмятин, охватывающих все слои раковины (Mironenko, 2016).
Особенно часто искривлению раковины аммонитов способствовали различные обрастатели, селившиеся на их поверхности, в первую очередь двустворчатые моллюски и черви-серпулиды (Larson, 2007; Keupp, 2012). Аммонит, будучи не в силах убрать обрастателя, был вынужден строить новый оборот раковины поверх поселенца, что приводило к искажению формы раковины, в некоторых случаях очень существенному. В некоторых случаях форма раковины становилась скафитоидной (Рис.5). Прикрепление крупного поселенца к одной из боковых сторон раковины могло к тому, что аммонит был вынужден изменять плоскость навивания раковины, чтобы компенсировать вес эпибионта (Keupp, 2012). В юрских отложениях Центральной России аномалии, вызванные обрастателями, встречаются не часто, за исключением верхнекелловейских отложений местонахождения Дубки, где на раковинах Quenstedtoceras поселенцы присутствуют в больших количествах (Сельцер, 2001; Larson, 2007; Сельцер, 2009; Сельцер, Косенко, 2015).

Рис. 5. Аномалии развития раковины, вызванные прикреплением эпибионтов. A, B – искривленные раковины Quenstedtoceras lamberti (верхний келловей, Дубки, Саратовская обл.), эпибионты скрыты под последним оборотом. Масштабнын отрезки 1 см для А и 3 см для В. С — асимметричная раковина Cardioceras cordatum (нижний оксфорд, Михайлов, Рязанская обл.). С1 — вид сбоку, эпибионт (двустворчатый моллюск) прикрепился в умбиликальной части раковины аммониты, его поверхность копирует форму ребер аммонита. Жилая камера деформирована посмертно. С2 — вид с вентра, киль отклонился в сторону из-за веса эпибионта. Масштабный отрезок — 5 мм.
Кроме болезней и паразитов аномальное развитие раковин, вероятно, могли вызывать и различные абиотические факторы. Среди вариантов «болезненных форм» раковин аммонитов есть forma aegra aptycha, характеризующаяся наличием дополнительной стенки внутри раковины и полостей, расположенных между этим дополнительным образованием и нормальной внешней стенкой. Двойная вентральная стенка иногда встречается у верхнеоксфордских Amoeboceras alternoides в Московской области, причем дополнительная внутренняя пластина даже несет небольшой киль (Рис.6A). Подобные аномалии описаны у современных наутилусов, развивавшихся в аквариумах в условиях неправильного питания или температурного режима (Keupp, 2012). Какие именно причины приводили к развитию подобной патологии у аммонитов неизвестно, но скорее всего это также были негативные условия среды — неподходящее питание или аномальные температуры, хотя влияние паразитов в этом случае также нельзя исключить.
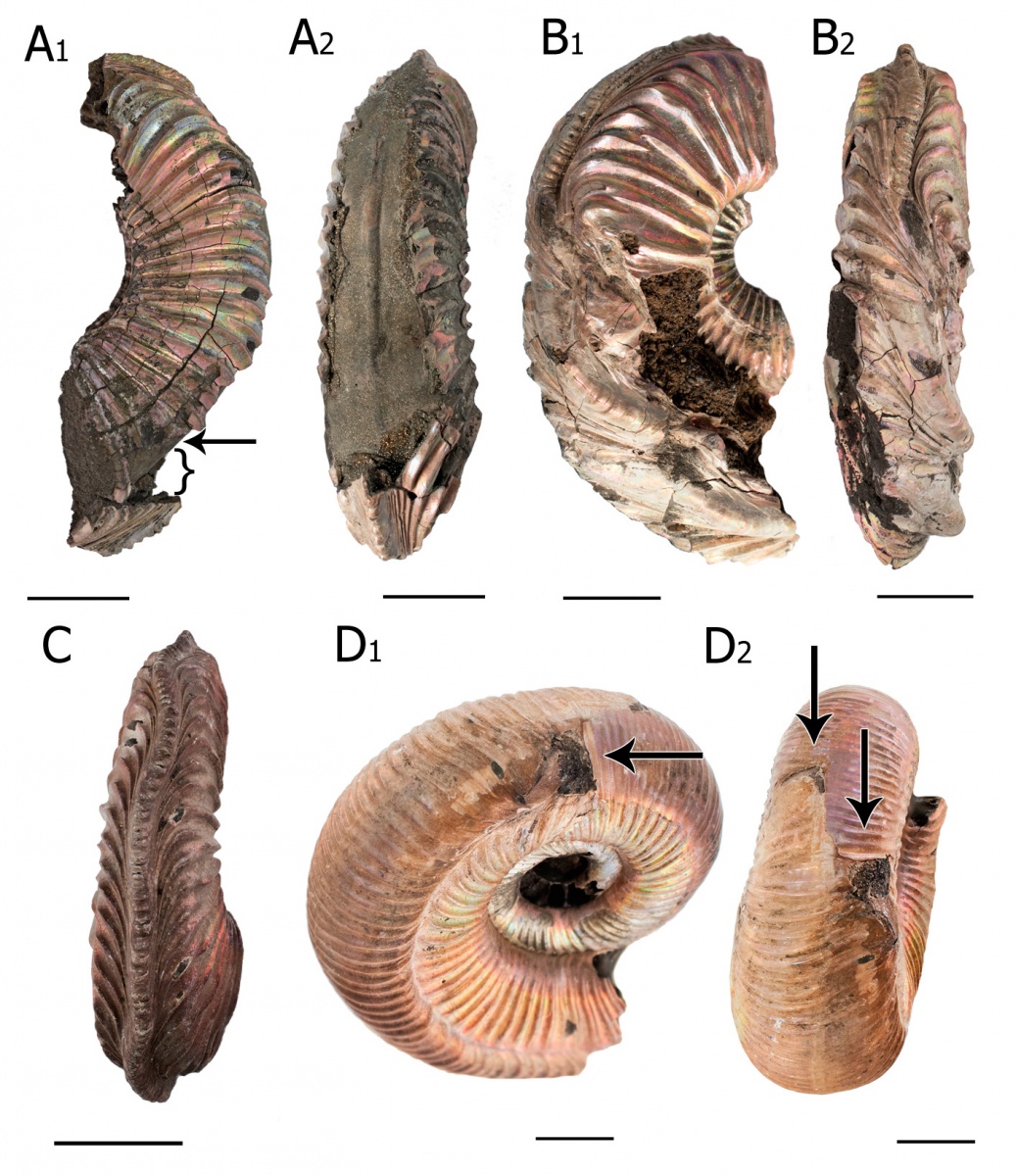
Рис. 6. Аномалии развития и прижизненные повреждения раковин аммонитов верхнего оксфорда, Марково, Московская обл. A – дополнительная внутренняя стенка раковины (forma aegra aptycha) в илой камере Amoeboceras alternoides. A1 – стрелка указывает на дополнительную стенку, фигурная скобка отмечает расстояние между этой пластиной и вентральной стенкой, несущей киль. A2 – обратите внимание на низкий киль на дополнительной пластине. B - Аномальное развитие вентральной части раковины (forma aegra chaotica) Amoeboceras alternoides. Скорее всего, причиной этой аномалии была серьезная травма, с последующим воспалением тканей мантии. С - Множественные изгибы киля (forma aegra undaticarinata) на раковине Amoeboceras alternoides. Никакие видимы следы повреждений не предшествуют этой аномалии. D -Крупное прижизненное повреждение с последующим шрамом на раковине Subdiscosphinctes sp. Масштабные отрезки — 1 см.
У современных наутилусов, живущих у дна среди рифов и скал и при этом плохо видящих, нередки характерные повреждения вентральной части жилой камеры — трещины и даже проломы, вызванные ударами о твердые предметы. Наутилусы успешно залечивают такие повреждения. Однако, на раковинах аммоноидей подобные травмы до сих пор не обнаружены.
Сравнение прижизненных повреждений и аномалий роста раковин аммонитов из различных местонахождений Центральной России.
В отложениях разного возраста на раковинах аммонитов встречаются различные прижизненные повреждения. Также существенно варьирует частота этих повреждений. Более того, даже в одновозрастных местонахождениях частота повреждений и их варианты могут существенно различаться. Изучение повреждений в отдельных местонахождениях позволяет понять, под давлением каких именно хищников или факторов окружающей среды находились аммониты и насколько велико было это давление. В этом разделе представлен обзор прижизненных повреждений раковин аммонитов из нескольких местонахождений средней и верхней юры.
Михайлов, средняя юра, средний и верхний келловей, зоны Erymnoceras coronatum и Peltoceras athleta.
В келловейских отложениях Михайлова встречаются арагонитовые раковины аммонитов хорошей сохранности. Как правило, их фрагмоконы изнутри выполнены кристаллами пирита, что способствует их прочности, в то время как не пиритизированные жилые камеры часто оказываются полностью раздавленными. Это существенно затрудняет извлечение и сохранение аммонитов с жилыми камерами, поэтому в основном здесь изучались повреждения, следы которых различимы на фрагмоконе.
Прижизненные повреждения на раковинах аммонитов в келловее Михайлова встречаются очень часто. Практически все они несут следы явного внешнего воздействия - скорее всего, это последствия атак хищников. В основном это различные повреждения края временного устья, довольно часто сопровождающиеся последующим искривлением формы раковины и иногда утерей скульптурных элементов. Утрата элементов скульптуры в результате повреждения мантийного края особенно характерна для представителей рода Kosmoceras, у которых в результате травмы может исчезать один или даже оба ряда шипов, такая посттравматическая аномалия именуется forma aegra calcar (Рис.3C,E).

Рис. 7. Прижизненные повреждения и аномалии на раковинах Binatisphinctes sp. (средний келловей, Михайлов, Рязанская область). A – Длинный шрам на раковине Binatisphinctes. Подобные шрамы отражают серьезное повреждение мантийного края. А1 — общий вид, А2 — увеличенный участок, обратите внимание на сдвиг шрама перед каждым временным параболическим устьем, это связано с формированием расширенного устья с параболическими бугорками (см. Radtke et al. 2016). B – Фрагмент раковины Binatisphinctes с одиночным параболическим бугорком. C - Фрагмент раковины Binatisphinctes со шрамом (forma aegra verticata) который начинается из параболического бугорка. Масштабные отрезки равны 1 см.
У представителей рода Binatisphinctes также присутствуют многочисленные шрамы, повреждения довольно часто концентрируются на параболических бугорках (Рис.7). Это подтверждает предположение о том, что параболические устья могли иметь защитное значение (Radtke et al. 2016). Кроме того, у Binatisphinctes не редки аномалии развития самих параболических бугорков, к примеру, появление одиночных, непарных бугорков. Такие аномалии редко сопровождаются шрамами или искривлением формы раковины, это заставляет предположить, что они имеют не травматическую природу, а являются следствием каких-то внутренних сбоев в функционировании мантийного края, формировавшего устья (о формировании параболических бугорков см. Radtke et al. 2016).
Binatisphinctes также являются рекордсменами по числу повреждений раковин не только среди келловейских аммонитов Михайлова, но и среди всех изученных автором юрских аммонитов Центральной России: более 16 процентов их раковин несут следы залеченных травм. На втором месте по числу травм (и в Михайлове и среди изученных аммонитов в целом) раковины Kosmoceras, среди них залеченные повреждения присутствуют на 15,54 % раковин (см. табл. 1). Интересно, что у обоих родов очень высок процент повреждений мантии, диагностирующийся по аномалиям на сформированной после травмы части раковины. Среди Kosmoceras у 50% поврежденных аммонитов есть следы повреждения мантии, среди Binatisphinctes их почти 43%.
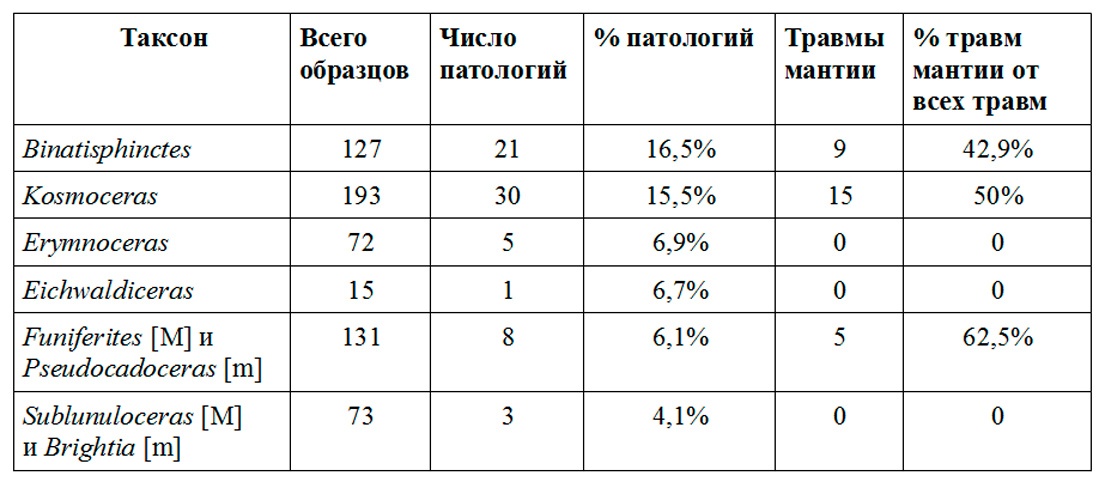
Таблица 1
Меньше всего прижизненных повреждений наблюдается на оксиконических раковинах диморфной пары Brightia [m] - Sublunuloceras [M] – лишь 4,1% изученных раковин несут следы залеченных травм, причем ни в одном из этих случаев, насколько можно судить по раковине, не был поврежден край мантии. Это может свидетельствовать о том, что оксиконические раковины реже других подвергались атакам хищников или избегали нападения благодаря своей скорости и манёвренности. Однако, с другой стороны, можно предположить, что сильные повреждения мантийного края могли быть летальными для этих аммонитов, поскольку значительное искривление раковины наверняка существенно снижало бы их манёвренность и подвижность, а это могло быть критическим для выживания особи. На двух раковинах Brightia сохранившихся с жилыми камерами были обнаружены смертельные вентральные укусы (Рис.4). К сожалению, пока находок этих аммонитов с жилыми камерами слишком мало для каких-либо выводов.
Также довольно низкий процент повреждений демонстрируют аммониты Erymnoceras: у этих аммонитов подобные аномалии отмечены у примерно 7% изученных экземпляров, причем, среди них тоже не найдено ни одной травмы, приведшей к существенному искривлению раковины.
Необычная ситуация наблюдается в диморфной паре Funiferites [M] - Pseudocadoceras [m]. В целом поврежденных раковин среди них не много — чуть больше 6%, но среди них больше половины (62,5%) имеют признаки повреждения мантии, причем среди изученных раковин Pseudocadoceras абсолютно все травмы сопровождаются серьезным искривлением последующей части раковины. Интересно, что наблюдающиеся у дискоконических Pseudocadoceras характерные повреждения в виде разрушения вентральной стороны и одной из латеральных стенок раковины также встречаются у дискоконических верхневолжских Craspedites. Такое сходство повреждений при сходной форме раковины может свидетельствовать о близком образе жизни этих аммонитов. Так как такие травмы интерпретируются как следы атак ракообразных (Keupp, 2012; Hoffmann, Keupp, 2015), можно предполагать, что и Pseudocadoceras и Craspedites обитали неподалеку от дна, где встречи с ракообразными были наиболее вероятны.
Еще один вариант травм, который может быть связан с атаками ракообразных — симметричные повреждения обеих латеральных стенок, при которых вентральная сторона остается нетронутой. Для нанесения такого повреждения хищник должен был хватать аммонита спереди либо одним тонким орудием, либо двумя — за каждую стенку раковины. Челюстью или клювом нанести такое повреждением практически невозможно, а вот клешни ракообразных в данном случае хорошо подходят на роль вероятного орудия атаки. Такие травмы редки среди всех аммонитов, в келловее Михайлова они отмечены у Erymnoceras и Pseudocadoceras.
Несколько экземпляров Funiferites и Erymnoceras имеют повреждения в виде полукруглых выбоин, очень похожих на следы, которые оставляют костистые рыбы на современных наутилусах (Рис.1B,C). Обычно такие повреждения на аммонитах интерпретируются как следы зубов рыб семейства Semionotidae (Keupp, 2012). В некоторых случаях эти повреждения имеют довольно значительные размеры, однако не сопровождаются возникновением шрамов.
Несколько лет назад статистика по прижизненным повреждениям аммонитов из келловея Михайлова была опубликована Г. Койпом (Keupp, 2012, табл.3, c.77). Согласно этим данным, среди Kosmoceras следы повреждений присутствуют на 13,6% раковин, среди Binatisphinctes на 13,2% и среди Hecticoceras (Brightia и Sublunuloceras в данной публикации) их 8,7%. Это вполне согласуется с подсчётами, полученными автором, хотя процент травмированных раковин Brightia и Sublunuloceras в сборах автора все-таки значительно меньше. Данные по тем же таксонам из других местонахождений в Западной Европе (Keupp, Ilg 1992; Keupp, 2012) свидетельствуют о том, что в Михайлове процент поврежденных раковин существенно выше, чем в других регионах. Столь большое количество травмированных раковин и значительная доля мантийных травм среди них говорят о том, что аммониты в этой экосистеме находились под значительным прессингом хищников.
Дубки, Средняя юра, верхний келловей, зона Quenstedtoceras lamberti
В Дубках в глинах зоны Quenstedtoceras lamberti встречаются многочисленные арагонитовые раковины аммонитов хорошей сохранности. Как и в келловейских отложениях Михайлова, их фрагмоконы пиритизированы и хорошо сохраняются, в то время как жилые камеры оказываются раздавленными и редко пригодны для изучения. Среди аммонитов здесь численно доминируют представители рода Quenstedtoceras и близкие к ним Vertumniceras [m] и Eboraciceras [M], составляющие вместе почти 90% находок. И именно Quenstedtoceras лидируют по числу различных палеопатологий. Явные травмы, в основном повреждения края временного устья, обнаружены у 14% Quenstedtoceras, правда, травм мантии среди них немного — 7,7% от числа всех повреждений. Сходные данные по прижизненным повреждениям Quenstedtoceras из Дубков приводятся Н.Ларсоном — 167 изученных им экземпляров из 1000 (16,7%) несли следы прижизненных повреждений (Larson, 2007).

Рис. 8. Раковина Quenstedtoceras lamberti (верхний келловей, Дубки, Саратовская обл.) с множественными прижизненными повреждениями (отмечены стрелками). Масштабный отрезок 1 см.
Некоторые из раковин Quenstedtoceras, изученных автором, имеют сразу несколько прижизненных повреждений, полученных на разных стадиях роста (Рис.8). Это может свидетельствовать о том, что искривление формы раковины вследствие травмы повышало уязвимость или заметность аммонита для хищников, но также возможно, что в некоторые периоды или сезоны численность хищников существенно возрастала и жившие в это время аммониты подвергались более частым атакам, чем остальные.
У некоторых макроконхов Quenstedtoceras часть повреждений приурочена к хорошо выраженному временному устью в начале взрослой гладкой стадии, незадолго до формирования конечной жилой камеры. Можно предположить, что это временное устье соответствует какой-то долговременной задержке роста, в течение которой аммониты могли получать незначительные травмы мантийного края, проявляющиеся после продолжения роста. Возможно, что эта задержка роста была связана с сезонными колебаниями температуры или количества пищи.
Также раковины Quenstedtoceras известны большим количеством аномалий, вызванных прикреплением эпибионтов к поверхности их раковин (Сельцер, 2001; Larson, 2007; Сельцер, 2009). Ларсон (2007) указывает, что почти 65% изученных им раковин Quenstedtoceras из Дубков несли следы различных палеопатологий, не связанных с воздействием хищников - это различные искривления раковины и горбы-«протуберанцы». (Рис.5A,B) Среди изученных автором Quenstedtoceras лишь 5% имеют искривления, вызванные обрастателями, однако и это очень большой процент по сравнению с остальными юрскими аммонитами Центральной России, так как обычно доля раковин с поселенцами значительно меньше 1%. Интересно, что на раковинах других таксонов, встречающихся в Дубках, поселенцы практически отсутствуют. Сельцер (2009) полагает, что это может быть связано с придонным образом жизни Quenstedtoceras и высокой концентрацией личинок двустворчатых моллюсков у дна, однако повреждения, связанные с атаками ракообразных на раковинах Quenstedtoceras не обнаружены, что ставит их придонный образ жизни под сомнение.
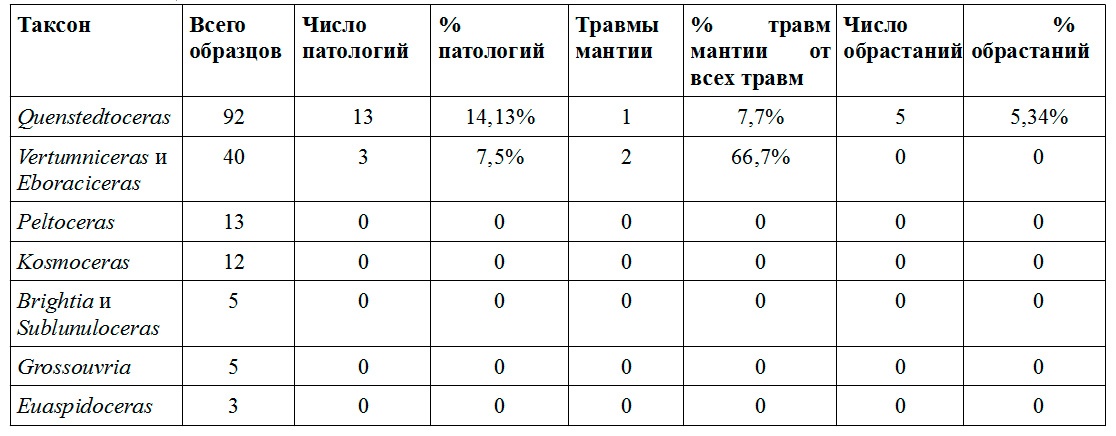
Таблица 2
У остальных таксонов аммонитов из Дубков прижизненные повреждения редки (см. табл. 2). Согласно данным Ларсона (2007), среди изученных им 100 образцов различных таксонов из Дубков (за исключением Quenstedtoceras) травмы и аномалии наблюдались на 89 раковинах. Однако, Сельцер (2001) сообщает, что в целом среди всех аммонитов Дубков (очевидно, включая и Quenstedtoceras) прижизненные повреждения были встречены на 10-12% раковин. Эти данные и редкость прижизненных повреждений и аномалий в изученных автором сборах (хотя выборка в данном случае невелика) заставляет предположить, что использовавшаяся Ларсоном коллекция могла изначально специально собираться из аномальных раковин.
Михайлов, верхняя юра, нижний оксфорд, зона Cardioceras cordatum
Сохранность аммонитов в нижнем оксфорде Михайлова практически не отличается от сохранности в описанных выше келловейских слоях того же местонахождения. Но по сравнению со средним и верхнем келловеем в нижнем оксфорде Михайлова аммониты с прижизненными повреждениями встречаются значительно реже. Если в келловее у некоторых таксонов до 16% всех раковин несут на себе следы залеченных повреждений, то в нижнем оксфорде число травмированных раковин не превышает 7% (см. табл. 3). Среди представителей рода Cardioceras только 6,23% раковин имеют следы повреждений, в 25% случаев повреждение сопровождается формированием шрамов или утерей киля.
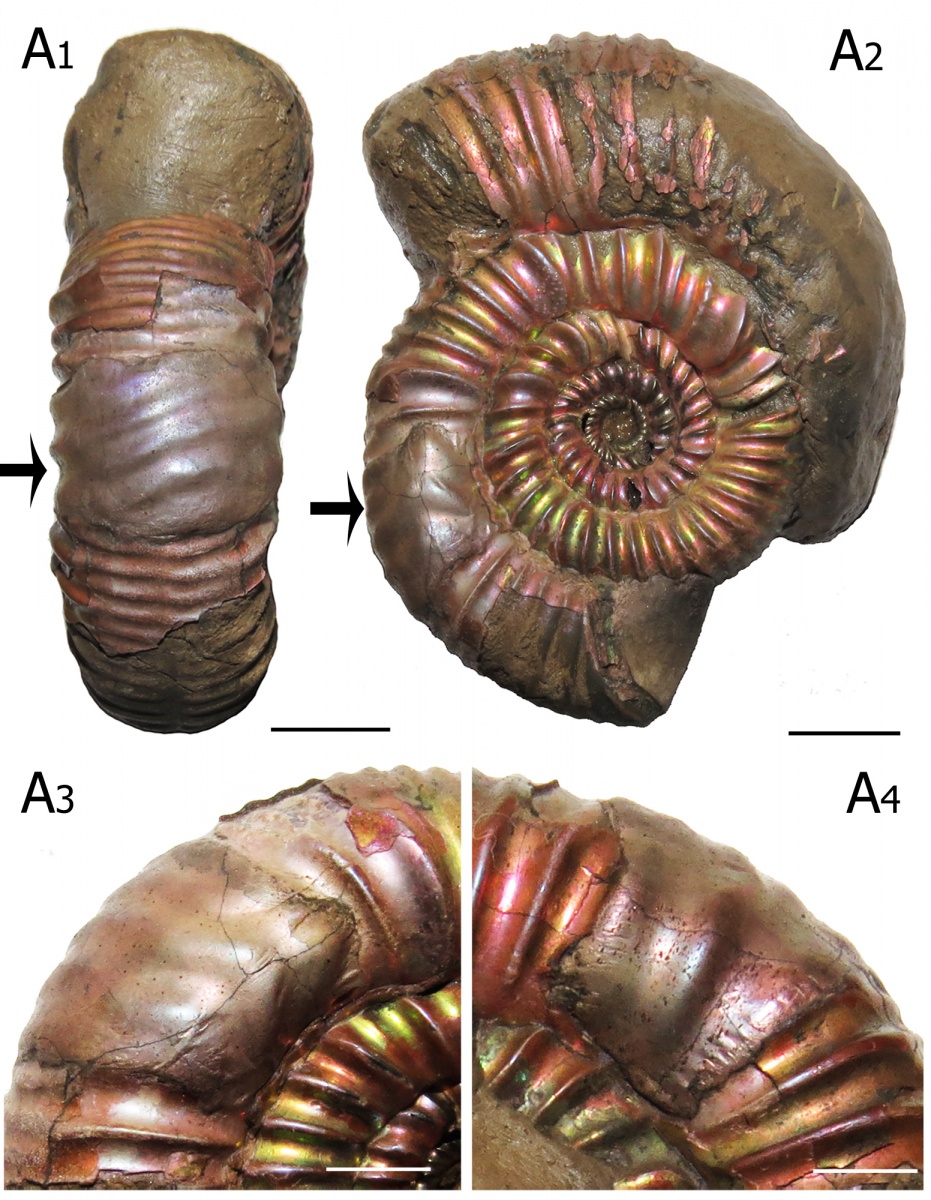
Рис. 9. Раковина аммонита Properisphinctes sp. (нижний оксфорд, Михайлов, Рязанская область) с залеченным отверстием стенки жилой камеры (forma aegra fenestra). В отличие от большинства прижизненных повреждений, связанных с травмами устьевого края, forma aegra fenestra представляет собой залеченный вентральный укус. А1 - вид с вентра, А2 - вид сбоку, А3 и А4 - вид с двух латеральных сторон. Длина масштабных отрезков на А1 и А2 = 1 см, на А3 и А4 = 5 мм.
Среди представителей рода Properisphinctes, встречающихся значительно реже, чем кардиоцератиды, была обнаружена лишь одна раковина с залеченным повреждением (что составляет 6,67% от изученных образцов), хотя и очень необычным (Рис.9). Это повреждение относится к так называемой forma aegra fenestra, это не повреждение устьевого края, а пролом в стенке раковины на некотором расстоянии позади устья. Причем, в случае данного Properisphinctes, это довольно обширное повреждение, являющееся по сути залеченным вентральным укусом (Keupp, 2006). Такие же залеченные вентральные укусы известны на раковинах верхнеоксфордских Perisphinctes из местонахождения Михаленино на р.Унже. В обоих случаях перисфинктиды с длинными жилыми камерами (около одного оборота) обитали вместе со значительно более многочисленными кардиоцератидами, обладавшими существенно более короткими жилыми камерами, имеющими длину от половины до двух третей оборота (это Cardioceras в раннем оксфорде и Amoeboceras в позднем). Из-за разницы в длине жилой камеры вентральный укус, нанесенный на одинаковом расстоянии от устья у кардиоцератид приходился на критически важную для моллюска область прикрепления мускулатуры в задней части жилой камеры, а у перисфинктид — лишь в среднюю часть жилой камеры вне мускульных отпечатков. Можно предположить, что такие залеченные вентральные укусы у перисфинктид - это результат ошибки хищников, атаковавших аммонитов с длинной жилой камерой на том же расстоянии от устья, на котором они привыкли пробивать раковины значительно более многочисленных кардиоцератид.
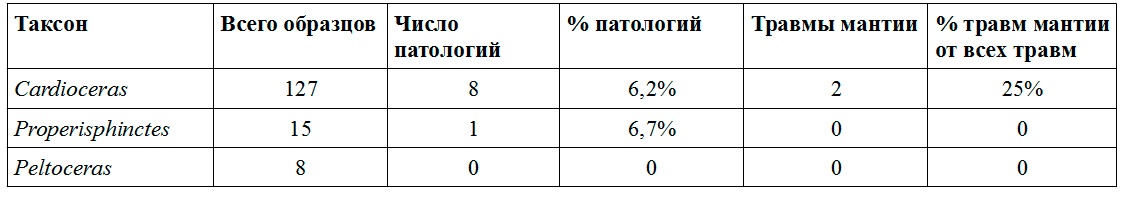
Таблица 3
Интересно, что изученные автором микроконхи и ювенильные макроконхи рода Peltoceras из нижнего оксфорда Михайлова не несут никаких следов травм, хотя их скульптура представлена лишь ребрами. Шипы, которые могли иметь защитное значение, появляются у макроконхов только на взрослой стадии развития раковины. Возможно, что эти аммониты каким-то образом избегали внимания хищников до тех пор, пока не достигали больших размеров. Впрочем, изученная автором выборка слишком мала для достоверных выводов.
Марково, верхняя юра, верхний оксфорд, зона Amoeboceras alternoides
Раковины аммонитов верхнеоксфордских слоях в Марково отличаются очень хорошей сохранностью. Сохраняется не только арагонитовая раковина, но нередко и морщинистый слой (Mironenko, 2015). Фрагмоконы, как и в Михайлове и Дубках, заполнены пиритом, однако в Марково, в отличие от большинства других местонахождений пиритизированных раковин аммонитов, довольно часто сохраняются пиритизированные жилые камеры.
Аммонитовая фауна в верхнеоксфордских слоях зоны Amoeboceras alternoides в Марково представлена лишь двумя родами: Amoeboceras и Subdiscosphinctes. Залеченные прижизненные повреждения на раковинах Amoeboceras в этом местонахождении встречаются довольно редко: они найдены на 5,61% изученных раковин, в 13,5% случаев повреждение, по-видимому, затронуло мантию (см. табл. 4). Однако, повреждения и аномалии у Amoeboceras alternoides очень разнообразны. Чаще всего встречается довольно специфический вид травмы: одна из латеральных стенок временного устья оказывается выломанной на довольно большую глубину, при этом ни вентральная сторона, несущая зазубренный киль, ни другая стенка повреждений не имеют (Рис. 1А). Из 14 таких травм 11 приходятся на правую сторону раковины и только 3 — на левую. Это первый случай обнаружения явной асимметрии в расположении прижизненных повреждениях у аммонитов (см. Kerr, Kelley, 2015). Изломанный край поврежденного участка наводит на мысль, что эти повреждения могли быть нанесены рыбами.

Рис. 10. Две раковины Amoeboceras alternoides (верхний оксфорд, Марково, Московская обл.) без киля на вентральной стороне (forma aegra circumdata). Форма ребер на таких раковинах отличается от типичной для данного вида: они изгибаются назад у вентра. Отсутствие киля и аномальная форма ребер делают определение подобных раковин сложной задачей. Длина масштабных отрезков 5 мм.
У двух изученных раковин Amoeboceras полностью отсутствует киль, а рёбра, выгнутые назад сильнее, чем у нормальных раковины, просто проходят через вентральную сторону (Рис.10). Подобная аномалия именуется forma aegra circumdata. Хотя на исследованных автором образцах место возникновения аномалии скрыто под более поздними оборотами, скорее всего, она связана с серьезным повреждением вентральной части мантии на ранних стадиях онтогенеза. Отсутствие киля и связанный с ним нетипичный изгиб ребер придают аномальным раковинам крайне необычный вид, несколько напоминающий представителей рода Peltoceras.
Две раковины Amoeboceras alternoides демонстрируют крайне редкий тип аномалии развития раковины, так называемую forma aegra aptycha (Рис.6A). Эта аномалия характеризуется формированием двойной стенки раковины с полостью внутри. Интересно, что у обоих Amoeboceras аномалия обнаружена в конечной жилой камере. Как упоминалось выше, подобные двойные стенки раковины описаны у современных наутилусов, развивавшихся в аквариумах в условиях неправильного питания или температурного режима. Какие именно причины приводили к формированию подобной аномалии у аммонитов неизвестно, но скорее всего, это также были либо негативные условия среды, либо деятельность паразитов.
Также у Amoeboceras alternoides встречается еще один необычный вид аномалии: forma aegra undaticarinata (Рис.6C), представляющая собой незначительное, но заметное виляние киля из стороны в сторону (Heller, 1964; Keupp, 2012). В отличие от искривления киля, связанного с тяжёлой травмой вентральной области (forma aegra chaotica, также встречающейся у Amoeboceras, см. Рис.6B) аномалия forma aegra undaticarinata часто возникает без связи с каким-либо заметным повреждением. Такая картина наблюдается и в Марково: 12% всех Amoeboceras alternoides в этом местонахождении имеют искривленный киль, но не несут никаких признаков повреждения, предшествующих этому искривлению. Точно такое же искривление киля без видимой причины описано у плинсбахских родов в Германии, таких как Amaltheus и Pleuroceras, имеющих сходную с кардиоцератидами форму раковины (Heller, 1964; Keupp, 2012). Некоторые исследователи считают, что причиной этой аномалии могла быть паразитарная инфекция, поражавшая край мантии (De Baets et al. 2015). Это, однако, представляется маловероятным, так как искривление киля практически у всех изученных аммонитов прекращается также внезапно, как и появляется, и такие аммониты не демонстрируют каких-либо признаков угнетения роста. Более того, искривление киля без видимого внешнего воздействия характерно для аммонитов разного геологического возраста и разных регионов (Amoeboceras, Amaltheus, Pleuroceras), имевших киль, в то время как не имевшие киля таксоны, жившие с ними по соседству, не демонстрируют никаких аномалий вентральной части раковины.
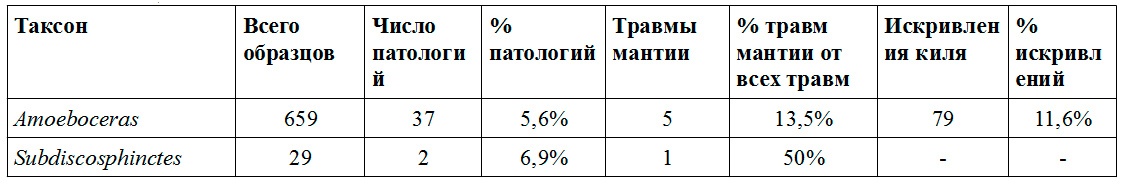
Таблица 4
Представляется более вероятным, что периодическое искривление киля могло быть связано с чисто механическими сбоями в процессе роста раковины, обладавшей на всех стадиях развития длинным и узким вентральным выступом — ростром: возможно, что при резких маневрах аммонита мантия и периостракум на самом кончике ростра могли несколько смещаться в сторону, что и приводило к искривлению растущей раковины.
Аммониты Subdiscosphinctes в Марково встречаются значительно реже, чем Amoeboceras. Автором изучено лишь 29 раковин представителей этого рода (по сравнению с более чем шестью сотнями Amoeboceras), при этом на двух образцах были найдены залеченные повреждения. В одном случае повреждение было очень значительным и сопровождалось появлением заметного шрама (Рис.6D). К сожалению, в отличие от Amoeboceras, раковины которых (за исключением крупных макроконхов) сохраняются в Марково практически целиком, часто даже с пиритизированными жилыми камерами, у Subdiscosphinctes, имевших значительно более крупные размеры, сохраняются только начальные обороты. Такая сохранность не позволяет объективно оценить уровень травмированности их раковин.
Палеоэкологические выводы
Сравнение залеченных прижизненных повреждений на раковинах аммонитов из разных местонахождений средней-верхней юры Центральной России показывает, что в разное время аммониты сталкивались с различными хищниками, паразитами и обрастателями и что давление различных неблагоприятных факторов на разные популяции аммонитов было неодинаковым.
Максимальным давление хищников на популяции аммонитов было во второй половине келловея на территории современной Рязанской области (Михайлов). Процент аммонитов с прижизненными повреждениями, связанными с атаками хищников, в келловее Михайлова существенно выше, чем в одновозрастных разрезах Западной Европы (Keupp, Ilg, 1992; Keupp, 2012) и в разрезах другого возраста в Центральной России. Особенно сильно в конце келловея страдали от нападений аммониты родов Kosmoceras и Binatisphinctes. Возможно, что именно это стало причиной возникновения в позднем келловее нового подсемейства Aspidoceratinae (предками которого, по-видимому, были Binatisphinctes), в котором раковины макроконхов были защищены шипами.
В терминальном келловее Дубков (Саратовская область) раковины с повреждениями, оставленными хищниками, тоже весьма многочисленны, но все-таки встречаются несколько реже, чем в Михайлове. Интересно, что здесь жертвами атак становились в основном кардиоцератиды, а не космоцератиды и перисфинктиды, как в Михайлове. Возможно, что это связано с численным преобладанием кардиоцератид в этих слоях (около 90% раковин аммонитов, найденных в Дубках, относятся к этому семейству). Благодаря их высокой численности данные по кардиоцератидам существенно полнее, чем по более редким таксонам, кроме того, вполне вероятно, что и хищникам было проще охотиться на доминирующие в экосистеме виды аммонитов, чем на редко встречающиеся. Также многие раковины рода Quenstedtoceras имеют различные аномалии роста, как связанные с прикреплением обрастателей к наружной поверхности их раковин, так и, возможно, с активностью паразитов.
В раннем оксфорде на территории современной Рязанской области, судя по находкам в Михайлове, давление хищников на аммонитов резко снизилось по сравнению келловеем. В среднем в келловее Михайлова и Дубков около 10% всех раковин имеют те или иные патологии, в нижнем оксфорде Михайлова поврежденных или аномальных раковин около 5%. Правда, существенно (до трех родов) снизилось и разнообразие аммонитов. Также очень невелик процент поврежденных раковин в верхнем оксфорде (разрез Марково в Московской области), где совместно встречаются лишь два рода аммонитов. О довольно низком уровне давления хищников говорят и сохраняющиеся в Марково жилые камеры аммонитов: на них практически не встречаются повреждения, которые можно было бы интерпретировать как смертельные для аммонита. Однако, здесь у представителей рода Amoeboceras встречаются различные редкие аномалии развития раковин, как посттравматического, так и неустановленного характера. Особенно интересно периодическое искривление киля, практически на каждом десятом образце. Хотя подобные искривления часто связывают с деятельностью паразитов, нельзя исключить, что причины были исключительно в механических трудностях строительства очень длинного и тонкого устьевого выроста-ростра.
Сравнение палеопатологий, обнаруженных на раковинах различных аммонитов, позволяет выявить таксоны, имеющие сходные травмы. Так, очень сходные между собой прижизненные повреждения, интерпретируемые как следы атак рыб, встречаются на раковинах Erymnoceras и Funiferites из келловея Михайлова. Раковины Pseudocadoceras (микроконхи, составляющие диморфную пару с Funiferites) и Erymnoceras имеют повреждения, которые могут быть связаны с атаками ракообразных. При этом точечные укусы, часто встречающиеся на раковинах Kosmoceras и Binatisphinctes, у Erymnoceras и Funiferites практически не наблюдаются. Исходя из всего этого, представляется вполне возможным, что аммониты родов Erymnoceras и Funiferites занимали сходные экологические ниши. Наличие следов атак ракообразных говорит о том, что областью обитания этих аммонитов были, скорее всего, придонные слои воды.
Большое количество травм, связанных с атаками хищников, на раковинах Kosmoceras и Binatisphinctes свидетельствует в пользу того, что эти аммониты, по всей видимости, обитали в наиболее конкурентных и опасных условиях, скорее всего недалеко от поверхности. К сожалению, довольно сложно сказать, какие именно хищники ответственны за большую часть травм у этих аммонитов. Однако, серьезные повреждения мантии, часто сочетающиеся с относительно некрупными повреждениями раковины, свидетельствуют в пользу хищников, способных очень быстро атаковать аммонитов до того, как они успевали втянуться в раковину.
На раковинах оксфордских кардиоцератид (Cardioceras и Amoeboceras) такие травмы встречаются существенно реже. Позднеоксфордские Amoeboceras чаще всего страдали от укусов рыб, однако, эти укусы не идентичны повреждениям на келловейских аммонитах, вероятно, их оставляли другие рыбы. При этом, данные хищники крайне редко задевали мантию аммонита, несмотря на относительно короткие жилые камеры Amoeboceras и довольно значительные размеры повреждений на раковинах.
Отдельные симметричные травмы боковых стенок, связанные, скорее всего, с атаками ракообразных, встречены на отдельных экземплярах оксфордских кардиоцератид. Однако, единичные находки, в отличие от многочисленных повреждений такого рода, не позволяют делать выводы о месте обитания этих аммонитов — хотя ракообразные, скорее всего, жили у дна, аммониты могли обитать в толще воды и получить травмы, случайно оказавшись в придонном слое. При этом возможность того, что аммониты сталкивались с ракообразными во время откладки яиц (Сельцер, 2001) можно полностью исключить, так как все подобные повреждения были получены аммонитами задолго до завершения роста и созревания. Кроме того, яйцеживорождение, обнаруженное недавно у аптских аммонитов (Mironenko, Rogov, 2016), скорее всего было свойственно всем аммонитам и никаких кладок у них не было.
Наличие заживших вентральных укусов (forma aegra fenestra) на раковинах оксфордских пересфинктид свидетельствует о том, что хищники иногда ошибочно атаковали их, принимая за кардиоцератид со значительно более короткими жилыми камерами. Правда, сложно сказать, с чем связаны эти ошибки — с недостаточным опытом самих хищников, с относительной редкостью перисфинктид или с тем, что эти аммониты могли обитать на других глубинах или иметь другие суточные циклы по сравнению с кардиоцератидами и реже сталкиваться с хищниками, охотившимися на кардиоцератид.
Благодарности
Автор благодарен Ирине Смуровой (Москва) за изготовление фотографий к этой публикации. Автор признателен Михаилу Рогову (ГИН РАН) за ценные замечания и идеи, позволившие существенно улучшить эту публикацию. Работа выполнена по теме ГИН РАН № 0135-2014-0064 при поддержке гранта РФФИ № 05-15-06183.
Литература
Несис К.Н. Головоногие: умные и стремительные (Истории из частной и семейной жизни кальмаров, каракатиц, осьминогов, а также наутилуса помпилиуса). М.: Издательство «Октопус”. 2005.
Сельцер В.Б. Об аномальных раковинах келловейских аммонитов // Труды НИИ Геологии СГУ. Новая серия. Саратов. 2001. Т.8. С. 29-45.
Сельцер В.Б. Аномальные фрагмоконы позднекелловейских кардиоцератид (Ammonoidea) // Леонова Т.Б., Барсков И.С., Митта В.В. (ред.) Современные проблемы изучения головоногих моллюсков. Морфология, систематика, эволюция, экология и биостратиграфия. (Москва, 2–4 апреля 2009 г.). М.: ПИН РАН, 2009. С.50-51.
Сельцер В.Б., Косенко И.Н. Эпиойкия устрицы Liostrea roemeri на аммонитах // Современные проблемы изучения головоногих моллюсков. Морфология, систематика, эволюция, экология и биостратиграфия. М.: ПИН РАН. 2015. 4.P. 120-123.
Тесакова Е.М., Шурупова Я.А., Устинова М.А. Стратиграфия келловея и нижнего оксфорда разреза Михайловцемент // Труды ГИН РАН. Вып. 615. С. 264-301.
Andrew C., Howe P., Paul C. R., Donovan S.K. Fatally bitten ammonites from the lower Lias Group (Lower Jurassic) of Lyme Regis, Dorset. // Proceedings of the Yorkshire Geological Society. 2010. Vol. 58. no.2. P. 81-94.
Dagys A. S., Weitschat W. Ammonoid jaws from the Boreal Triassic realm (Svalbard and Siberia) // Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg. 1988. Hft.67. P. 53-71.
De Baets K., Klug C., Korn D. Devonian pearls and ammonoid-endoparasite co-evolution // Acta Palaeontologica Polonica. 2011. Vol. 56. no.1. P. 159–180.
De Baets K., Keupp H., Klug C. Parasites of ammonoids // Ammonoid Paleobiology: From anatomy to ecology. Springer Netherlands, 2015. P. 837-875.
Engeser T., Keupp H. Phylogeny of the aptychi-posessing Neoammonoidea (Aptychophora nov., Cephalopoda) // Lethaia. 2002. Vol. 34. P.79–96.
Hengsbach R. Ammonoid pathology // In: Landman N.H., Tanabe K., Davis R.A. (eds.) Ammonoid paleobiology. New York: Plenum press, 1996. P.581-605.
Heller F. Neue Falle von Gehause-Mißbildungen bei Amaltheiden // Paläontologische Zeitschrift. 1964. Bd. 38. Hft. 3-4. S. 136-141.
Hölder H. Über Anomalien an jurassischen Ammoniten // Paläontologische Zeitschrift. 1956. Bd. 30. Hft. 1. P. 95-107.
Hoffmann R., Keupp H. Ammonoid paleopathology //Ammonoid Paleobiology: From anatomy to ecology. Springer Netherlands, 2015. P. 877-926.
Kerr J.P., Kelley P.H. Assessing the influence of escalation during the Mesozoic Marine Revolution: shell breakage and adaptation against enemies in Mesozoic ammonites // Palaeogeography, Palaeoclimatology, Palaeoecology. 2015. Vol. 440. P. 632-646.
Keupp H. Sublethal punctures in body chambers of Mesozoic ammonites (forma aegra fenestra n.f.), a tool to interpret synecological relationships, particularly predator-prey interactions. // Paläontologische Zeitschrift. 2006. Bd. 80. Hft. 2. P. 112-123
Keupp H. Atlas zur Paläopathologie der Cephalopoden // Berliner paläobiologische Abhandlungen. 2012. Bd. 12.S. 1–390
Keupp H., Ilg A. Paläopathologie der Ammonitenfauna aus dem Obercallovium der Normandie und ihre palökologische Interpretation // Berliner Geowissenschaftlische Abhhandlungen, Reiche E. 1992. Bd.3. S. 171-189.
Kiselev D., Rogov M., Glinskikh L., Guzhikov A., Pimenov M., Mikhailov A., Dzyuba O., Matveev A., Tesakova E. Integrated stratigraphy of the reference sections for the Callovian/Oxfordian boundary in European Russia // Volumina Jurassica. 2013. Vol. XI. P. 59-96.
Klompmaker A.A., Waljaard N.A., Fraaije R.H.B. Ventral bite marks in Mesozoic ammonoids // Palaeogeography, Palaeoclimatology, Palaeoecology. 2009. Vol. 280. P. 245-257.
Kröger B. On the ability of withdrawing of some Jurassic ammonoids // Abhandlungen der Geologischen Bundesanstalt. 2002. Bd. 57. P. 199–204.
Kruta I., Landman N., Rouget I., Cecca F., Tafforeau P. The role of ammonites in the Mesozoic marine food web revealed by jaw preservation // Science. 2011. Vol. 331. no.6013. P. 70-72.
Larson N. Deformities in the Late Callovian (Late Middle Jurassic) ammonite fauna from Saratov, Russia // In: Landman NH, Tanabe K, Mapes RH (eds) Cephalopods-present and past: new insights and fresh perspectives. New York: Springer,. 2007. P 344-374.
Makowski, H. Problem of sexual dimorphism in ammonites. // Palaeontologia Polonica. 1962. Vol.12. P. 1–92.
Mironenko A.A. Wrinkle layer and supracephalic attachment area: implications for ammonoid paleobiology. // Bulletin of Geosciences. 2015. Vol. 90. no.2. P. 389–416
Mironenko A.A. A new type of shell malformation caused by epizoans in Late Jurassic ammonites from Central Russia // Acta Palaeontologica Polonica. 2016. Vol. 61. P. 645–660.
Mironenko A.A., Rogov M.A. First direct evidence of ammonoid ovoviviparity // Lethaia. 2016. Vol. 49. No.2 P. 245-260.
Norman M.D., Paul D., Finn J., Tregenza T. First encounter with a live male blanket octopus: The world's most sexually size-dimorphic large animal // New Zealand Journal of Marine and Freshwater Research. 2002. Vol. 36. No.4. P.733-736.
Radtke G., Hoffmann R., Keupp H. Form and formation of flares and parabolae based on new observations of the internal shell structure in lytoceratid and perisphinctid ammonoids. // Acta Palaentologica Polonica. 2016. Vol. 61. P. 503–517.
Saunders W. B., Spinosa C., Davis L.E. Predation on Nautilus // Nautilus: The Biology and Paleobiology of a Living Fossil. Reprint with additions. Springer Netherlands. 2010. P. 201-212.
Wani R., Jenkins R. G., Mapes R.H. Preferential predatory peeling: ammonoid vs. nautiloid shells from the Upper Carboniferous of Texas, USA // Geobios. 2012. Vol. 45. P. 129-137.
Ward P. Shell sculpture as a defensive adaptation in ammonoids // Paleobiology. 1981. Vol. 7. no.1). P. 96-100.
Ключевые слова (тэги): прижизненные повреждения, ammonoid sublethal injuries
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 24 сентября 2017 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://ammonit.ru./text/2078.htm#306945">http://ammonit.ru./text/2078.htm#306945</a> |
|
|||||
Как интересно!!! | ||||||
|
комментарий 1 уровня |
Комментарий создан 24 сентября 2017 года в 18:35:54 |
|||||
 Добавить в избранное
Добавить в избранное 


|
<a href="http://ammonit.ru./text/2078.htm#306961">http://ammonit.ru./text/2078.htm#306961</a> |
|
|||||
Спасибо! Буду продолжать работу в этом направлении )) | ||||||
|
комментарий 2 уровня |
Комментарий создан 24 сентября 2017 года в 20:21:02 |
|||||

|
<a href="http://ammonit.ru./text/2078.htm#306946">http://ammonit.ru./text/2078.htm#306946</a> |
|
|||||
Александр, огромное спасибо за потрясающую (еще одну :)) публикацию! Такой труд! Очень интересно было читать и смотреть фото! Класс! :) | ||||||
|
комментарий 1 уровня |
Комментарий создан 24 сентября 2017 года в 18:41:12 |
|||||

|
<a href="http://ammonit.ru./text/2078.htm#306960">http://ammonit.ru./text/2078.htm#306960</a> |
|
|||||
Спасибо!!! | ||||||
|
комментарий 2 уровня |
Комментарий создан 24 сентября 2017 года в 20:20:35 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#306955">http://ammonit.ru./text/2078.htm#306955</a> |
|
|||||
Очень хорошо потрудился Александр! Снимаю шляпу :) на рис. 8 ошибка - Дубки Рязанская область | ||||||
|
комментарий 1 уровня |
Комментарий создан 24 сентября 2017 года в 19:59:57 |
|||||

|
<a href="http://ammonit.ru./text/2078.htm#306958">http://ammonit.ru./text/2078.htm#306958</a> |
|
|||||
Спасибо! К сожалению, в конце имела место некоторая спешка, из-за этого ошибки (( Поправил область. | ||||||
|
комментарий 2 уровня |
Комментарий создан 24 сентября 2017 года в 20:13:11 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#306999">http://ammonit.ru./text/2078.htm#306999</a> |
|
|||||
Александр.вопрос немного не по теме.
| ||||||
|
комментарий 1 уровня |
Комментарий создан 24 сентября 2017 года в 22:57:16 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#307000">http://ammonit.ru./text/2078.htm#307000</a> |
|
|||||
Все зависит от правил конкретного издания и от требований редакторов и рецензентов. Расположение аммонитов раньше было жестко стандартизированным, но сейчас, по-моему, за этим уже никто особенно не следит, кроме разве что каких-то совсем консервативных изданий. Вот например моя статья в серьезном журнале: Wrinkle layer and supracephalic attachment area: Implications for ammonoid paleobiology - там аммониты ориентированы совершенно по-разному. Иллюстрации обычно цветные в pdf и черно-белые в печати.
| ||||||
|
комментарий 2 уровня |
Комментарий создан 24 сентября 2017 года в 23:11:12 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#307006">http://ammonit.ru./text/2078.htm#307006</a> |
|
|||||
Спасибо. увидел.
| ||||||
|
комментарий 3 уровня |
Комментарий создан 24 сентября 2017 года в 23:17:53 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#307193">http://ammonit.ru./text/2078.htm#307193</a> |
|
|||||
Видимо, для этого и делают такие разночтния в оформлении :) | ||||||
|
комментарий 4 уровня |
Комментарий создан 26 сентября 2017 года в 18:36:40 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#307537">http://ammonit.ru./text/2078.htm#307537</a> |
|
|||||
Скажи спасибо, что по ГОСТу приходится ссылки оформлять )))
| ||||||
|
комментарий 3 уровня |
Комментарий создан 30 сентября 2017 года в 21:39:52 |
|||||

|
<a href="http://ammonit.ru./text/2078.htm#307038">http://ammonit.ru./text/2078.htm#307038</a> |
|
|||||
Интересные кривобочки) Даже не думала, что такие бывают) | ||||||
|
комментарий 1 уровня |
Комментарий создан 25 сентября 2017 года в 12:29:01 |
|||||

|
<a href="http://ammonit.ru./text/2078.htm#307062">http://ammonit.ru./text/2078.htm#307062</a> |
|
|||||
Интересная и нужная публикация!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 25 сентября 2017 года в 15:47:47 |
|||||

|
<a href="http://ammonit.ru./text/2078.htm#307068">http://ammonit.ru./text/2078.htm#307068</a> |
|
|||||
Спасибо! В литературе их много пилили, к примеру вот: Фото из публикации Larson N.L. 2007. Deformities in the Late Callovian (Late Middle Jurassic) Ammonite Fauna from Saratov, Russia. | ||||||
|
комментарий 2 уровня |
Комментарий создан 25 сентября 2017 года в 16:28:19 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#307113">http://ammonit.ru./text/2078.htm#307113</a> |
|
|||||
Замечательная публикация! По материалу и изложению хорошо видно, что данная работа планировалась, пополнялась материалом и воплощалась не один год! | ||||||
|
комментарий 1 уровня |
Комментарий создан 25 сентября 2017 года в 20:35:53 |
|||||

|
<a href="http://ammonit.ru./text/2078.htm#307192">http://ammonit.ru./text/2078.htm#307192</a> |
|
|||||
Спасибо! Да, я эту статистику с подсчетом аммонитов собирал несколько лет, а некоторые фото давно хотел опубликовать, но не было повода. | ||||||
|
комментарий 2 уровня |
Комментарий создан 26 сентября 2017 года в 18:36:10 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#307177">http://ammonit.ru./text/2078.htm#307177</a> |
|
|||||
Спасибо, за познавательную публикацию.
| ||||||
|
комментарий 1 уровня |
Комментарий создан 26 сентября 2017 года в 13:25:16 |
|||||

|
<a href="http://ammonit.ru./text/2078.htm#307191">http://ammonit.ru./text/2078.htm#307191</a> |
|
|||||
Спасибо! | ||||||
|
комментарий 2 уровня |
Комментарий создан 26 сентября 2017 года в 18:35:05 |
|||||
|
<a href="http://ammonit.ru./text/2078.htm#307218">http://ammonit.ru./text/2078.htm#307218</a> |
|
|||||
Бесспорно - превосходный научный труд! | ||||||
|
комментарий 1 уровня |
Комментарий создан 27 сентября 2017 года в 03:42:21 |
|||||
{kind=link}