|
|
 |
|
Анатомия и некоторые аспекты палеобиологии аммоноидей: современные данные
Автор: Alex
Рубрика: Палеонтологические публикации
Уважаемые читатели Аммонит.ру! В начале сентября вышла из печати книга Труды Геологического института РАН. Выпуск № 615: Юрские отложения юга Московской синеклизы и их фауна, подготовленная коллективом авторов из ГИНа, ПИНа и МГУ. С одной из публикаций этого сборника вы уже знакомы: Юрские морские рептилии Москвы и Подмосковья. В сборнике вышли две моих статьи, которые я, конечно, опубликую на Аммонит.ру. Ниже представлен текст одной из них, она называется Анатомия и некоторые аспекты палеобиологии аммоноидей: современные данные, в виде pdf-файла ее можно скачать на ResearchGate: Anatomy and some aspects of ammonoid palaeobiology current view. Эта публикация представляет собой обзор современных представлений об анатомии (и немного о палеобиологии) аммоноидей, в основном мезозойских аммонитов. Многие упомянутые здесь открытия связаны с находками, сделанными на территории юга Московской синеклизы и в целом в Центральной России.
----------------------------------------------
Анатомия и некоторые аспекты палеобиологии аммоноидей: современные данные
Мироненко А.А., Геологический институт РАН, Москва
Резюме
Аммоноидеи — подкласс головоногих моллюсков, существовавший в морях с конца девона до самого конца мела. Благодаря красоте и многообразию форм, раковины аммоноидей стали предметом коллекционирования, а быстрая эволюция и широкое распространение сделали их важнейшим инструментом биостратиграфии. Однако, несмотря на многочисленность раковин аммоноидей в палеозойских и мезозойских отложениях, на их популярность у любителей палеонтологии и их значение для стратиграфии, о самих аммоноидеях - об их внешнем облике, анатомии и образе жизни, до недавнего времени было известно очень и очень мало. Но в последние два десятилетия ситуация значительно изменилась.
Новые находки раковин аммоноидей хорошей сохранности пролили свет на строение мускульной системы аммоноидей и областей прикрепления мягких тканей к раковине, строение сифона, пищеварительной системы и челюстного аппарата. Детальное изучение эмбрионального развития современных головоногих и новейшие открытия в области эволюции всего класса цефалопод и позволило сделать выводы о строении тех частей мягкого тела аммоноидей, которые пока еще не обнаружены в ископаемом состоянии. В настоящее время можно считать доказанным, что аммоноидеи имели хорошо развитые глаза, сложную систему мускулов-ретракторов, мощную воронку и морщинистый слой, сходный с чёрным слоем раковины наутилид и игравшим важнейшую роль в плавании аммоноидей. Исходно аммоноидеи, как и другие головоногие, имели десять щупалец. Мягкие ткани их сифона были сходны с тканями сифонов современных наутилусов, хотя и отличались некоторыми деталями.
Данная публикация обобщает современные данные об анатомическом строении аммоноидей, в том числе базирующиеся на находках, сделанных автором на юрских разрезах Центральной России. Здесь также обсуждаются вопросы палеобиологии аммоноидей, такие как прижизненная ориентациях их раковин в воде, механизм плавания и особенности размножения.
Ключевые слова: аммоноидеи, аммониты, палеобиология, отпечатки мускулов, сифон, челюстной аппарат.
Введение
Изучение мягких тканей аммоноидей является сложной задачей из-за того, что они очень редко и плохо сохраняются в ископаемом состоянии. В отличие от ископаемых колеоидей, полностью сохранившиеся отпечатки тел и фосфатизированные «мумии» которых известны из мезозойских чёрных сланцев и плитчатых литографских известняков, ни одного относительно полного отпечатка тела аммоноидеи до сих пор не найдено. Отсутствие находок вовсе не говорит о том, что мягкие ткани аммоноидей чем-либо принципиально отличались от тканей других цефалопод: точно также отсутствуют находки тел ископаемых наутилоидей, включая предков современного наутилуса. Все дело в наружной раковине, которая препятствовала быстрому и полному захоронению тела в донном осадке и в которую тело могло втягиваться после смерти.
Поэтому для реконструкции внешнего вида и внутреннего строения аммоноидей приходится использовать не только палеонтологические находки, но и данные по строению других современных и вымерших головоногих моллюсков — наутилоидей и колеоидей. Особенно важное значение здесь имеет изучение эмбрионального развития современных головоногих, позволяющее отличить исходные черты этого класса от различных эволюционных приобретений, характеризующих ту или иную группу цефалопод.
Впрочем, в последние десятилетия изучение палеонтологических находок получило новый импульс благодаря современным технологиям, таким как сканирующая электронная микроскопия и трехмерная рентгеновская томография. Эти технологии позволили не только обнаружить новые детали строения аммоноидей, ранее скрывавшиеся от глаз исследователей, но и развеять некоторые заблуждения, возникшие из-за недостаточной разрешающей способности технологий, применявшихся в 50-80-хх годах ХХ века.
По современным представлениям, аммоноидеи не были ни своеобразными наутилусами, ни «колеоидеями в раковинах», как это иногда предполагалось ранее. Это был совершенно особенный подкласс головоногих моллюсков, хотя многие черты современных головоногих причудливым образом комбинировались в их строении.
В этой публикации дается краткий обзор современных представлений об анатомии и внешнем облике аммоноидей и в первую очередь юрских и меловых аммонитов.
Мускулатура и отпечатки мускулов
Фрагменты мягких тканей аммоноидей крайне редко сохраняются в ископаемом состоянии и в основном исследователям приходится работать только с раковинами этих моллюсков. Однако, задачу изучения строения мягкого тела аммонитов облегчает тот факт, что их раковины — это не только средство защиты от хищников и гидростатический аппарат, но и наружный скелет, служивший для прикрепления мышц и для опоры тканей. Изучение мест прикрепления мускулатуры к раковине (так называемых «отпечатков мускулов») позволяет получить подробное представление о мускульной системе аммоноидей. Отпечатки мускулов хорошо видны на ядрах жилых камер аммоноидей в тех случаях, когда раковина разрушена полностью или когда на ядре сохраняется лишь тонкий полупрозрачный внутренний слой раковины. Отпечатки мускулов описывались в палеонтологической литературе начиная с 19 века (Trautschold, 1871), однако, до недавнего времени полная картина расположения областей соединения мускулатуры с раковиной у мезозойских аммонитов оставалась непонятной.
Лучше всего на ядрах жилых камер сохраняются три области прикрепления мускулатуры: две на дорсо-латеральном перегибе умбиликальных стенок и одна в центре вентральной стороны, напротив сифона. Все они расположены недалеко от последней септы, хотя у некоторых гетероморфных аммонитов, к примеру у Audouliceras они могут быть существенно смещены вперёд (Doguzhaeva, Mapes, 2015). Такие отпечатки мускулов можно найти на ядрах раковин верхнекелловейских макроконхов Quenstedtoceras и Funiferites в карьере Пески, а также на ядрах верхнеоксфордских Amoeboceras, Mirosphinctes и Euaspidoceras в разрезе у д. Рыбаки (рис.1). Эти отпечатки мускулов являются, по-видимому, гомологичными областям прикрепления мускулов-ретракторов у современных наутилусов (Mironenko, 2015a). Хотя у наутилусов не три, как было у аммонитов, а лишь два отпечатка мускулов, расположенных на дорсо-латеральных перегибах у последней септы и они имеют очень крупные размеры, расположение этих отпечатков мускулов относительно заднего края мантии и их строение совершенно одинаковы у аммоноидей и современных наутилид.
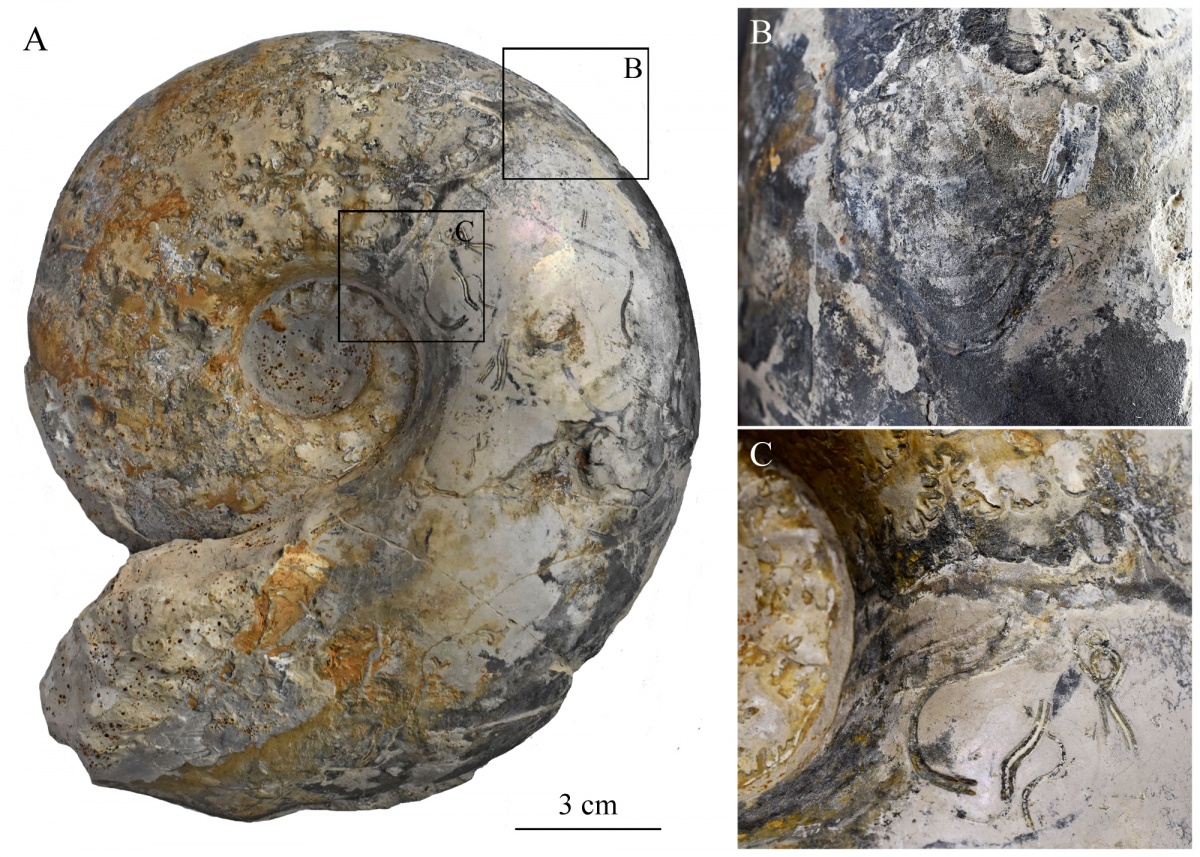
Рис. 1. Отпечатки мускулов на ядре раковины макроконха Quenstedtoceras из карьера Пески, Московская область, средняя юра, верхний келловей. А — общий вид образца, жилая камера короткая (меньше половины оборота). B — вентральный отпечаток мускула. C — дорсо-латеральный отпечаток мускула. Рядом видны трубки червей-серпулид, живших в жилой камере после смерти аммонита.
Мускулы-ретракторы головы, крепящиеся у наутилид к этим областям, имеют принципиальное значение для плавания наутилусов — именно они втягивают головной отдел моллюска в раковину, при этом голова работает как поршень, выжимая воду из мантийной полости (Несис, 2005). Вода с силой выталкивается через воронку, а моллюск движется в противоположную сторону. Малые размеры отпечатков мускулов аммонитов, по сравнению с отпечатками у наутилусов, в свое время дали многим исследователям основание предполагать, что аммониты были практически неспособны к активному плаванию (Kennedy, Cobban, 1976). Однако, в настоящее время изучение отпечатков мускулов различных представителей подкласса Nautiloidea показало, что мускульная система наутилусов уникальна среди головоногих моллюсков и ни у кого из наутилоидей, кроме представителей отряда Nautilida, такого «поршневого» механизма плавания не было.
При этом у аммоноидей мускульная система в целом оказалась устроена значительно сложнее, чем у наутилид. Начиная с 90-х годов 20 века появились публикации, посвященные находкам у аммонитов крупных латеральных (или вентро-латеральных) отпечатков мускулов (Догужаева, Кабанов, 1988; Doguzhaeva, Mutvei 1991, 1993, 1996). Эти отпечатки по своей структуре отличаются от отпечатков мускулов наутилусов и от упомянутых выше отпечатков мускулов аммоноидей. Так, расположенные недалеко от последней септы отпечатки мускулов и у аммоноидей и у наутилид образованы толстым слоем конхиолина, в то время как латеральные отпечатки мускулов аммонитов сформированы из дополнительных перламутрового и призматического слоев арагонита, то есть в этих областях стенка раковины состоит не из трех слоев, как обычно, а из пяти (Doguzhaeva, Mutvei, 1991). Кроме латеральных отпечатков мускулов была обнаружена еще одна структура: латеральные синусы, они по своей структуре были очень схожи с латеральными отпечатками, но, в отличие от них, очерченный синусами контур открыт в сторону устья (Doguzhaeva, Mutvei, 1996; Doguzhaeva, Mapes, 2015). Такая форма свидетельствовует о том, что местами прикрепления мускулов-ретракторов синусы быть не могли и их функция оставалась непонятной, кроме того, было совершенно не ясно как латеральные синусы соотносятся с латеральными мускулами.

Рис. 2. Латеральные отпечатки мускулов юрских аммонитов. A — Amoeboceras alternoides, верхняя юра, верхний оксфорд, Марково, Московская область. Латеральные отпечатки мускулов у аммонитов Amoeboceras ранее известны не были. B — Garniericeras catenulatum, верхняя юра, верхняя волга, Кунцево, Москва. На обоих образцах отпечатки мускулов выглядят как золотистые области в первой половине жилой камеры, но несколько различаются по форме: у Amoeboceras их ширина не очень велика, в то время как у Garniericeras они более широкие и сливаются на вентре.
Эти вопросы удалось разрешить благодаря изучению верхнеюрских аммонитов из семейства Craspeditidae уникальной сохранности, встреченных в верхневолжских отложениях Москвы и Московской области (Еганово, Карамышевская набережная и Кунцево), а также на р.Черемухе в Ярославской области. В этих отложениях на раковинах аммонитов Kachpurites и их потомков Garniericeras сохраняются одновременно и латеральные отпечатки мускулов и латеральные синусы, а также небольшие дорсо-латеральные и вентральные отпечатки (Mironenko, 2015a, Рис.2). В приустьевой части латеральных синусов этих аммонитов были обнаружены округло-треугольные области, служившие, по всей видимости, областями прикрепления мягких тканей (Mironenko, 2015a). И латеральные отпечатки мускулов и латеральные синусы всегда располагаются в вентро-латеральной области, ближе к вентральной стороне. Сравнение этих областей с областями прикрепления мягких тканей у современных колеоидей (Bizikov, 2002, 2008) дало основание для предположения, что латеральные отпечатки мускулов являются областями прикрепления ретракторов воронки. У наутилусов эти области в принципе отсутствуют, так как их ретракторы воронки крепятся к поверхности других мышц — ретракторов головы, а вот у колеоидей ретракторы воронки и области их прикрепления прекрасно развиты.
Расположение латеральных синусов и особенно субтреугольных структур в их приустьевой части говорит о том, что эти области могли использоваться для опоры мантийно-вороночных замыкателей (Mironenko, 2015 a). У наутилусов таких структур нет, а у современных колеоидей мантийно-вороночный замыкатель позволяет временно соединять воронку со стенкой мантии, что важно для быстрого плавания (Bizikov, 2008). У аммоноидей, по-видимому, для большей надежности соединения, мантийная часть этой структуры могла (скорее всего на необходимое время, а не постоянно) соединяться со стенкой раковины. Таким образом, мускульная система аммонитов была устроена намного сложнее, чем мускульная система наутилид и была во многом сходна с таковой у колеоидей. Наверняка плавали аммониты ничуть не хуже наутилусов, а скорее всего, намного лучше и быстрее (Doguzhaeva, Mutvei, 1993).
Воронка
Если предположения о функциях латеральных синусов и отпечатков мускулов верны, это означает, что аммоноидеи имели очень хорошо развитую воронку (Mironenko, 2015 a). В целом, наличие воронки у аммоноидей не вызывает сомнений, так как у всех их современных родственников, как относительно близких (колеоидей), так и дальних (наутилид) воронка есть (Klug, Lehmann, 2015). Оценить размеры воронки юрских аммонитов позволяет форма отверстия, образованного ушками многих микроконхов (Westermann, 1954, табл. 9, фиг. 1, табл. 22, фиг. 1, 2, табл. 28, фиг. 1, 8; Рис. 3 здесь). Судя по этим отверстиям, воронка должна была быть очень крупной: относительно размеров головного отдела она была крупнее, чем у большинства современных колеоидей, и в некоторых случаях занимала более трети высоты устья. При этом, как ни странно, художники-палеореконструкторы, изображающие аммоноидей, довольно часто забывают о воронке и либо не рисуют вовсе, либо изображают как маленький «довесок» где-то под щупальцами.
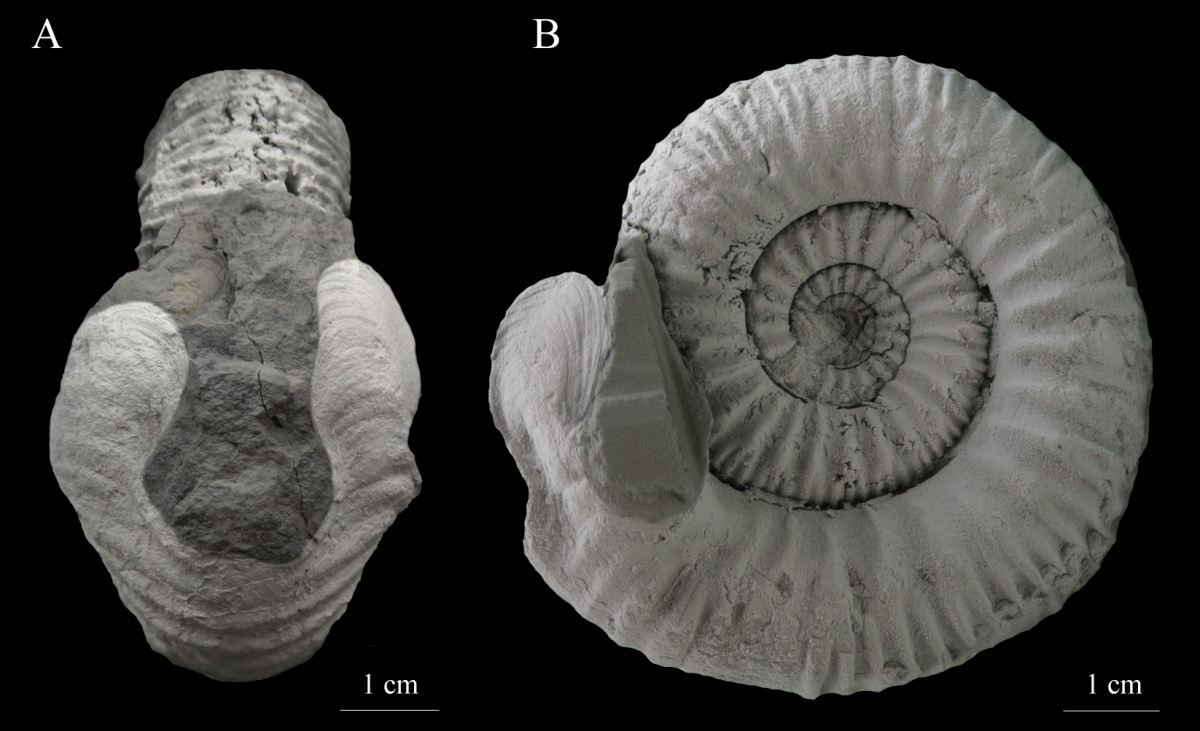
Рис. 3. Латеральные выросты (ушки) на устье микроконха Normannites, средняя юра, байос, Дагестан. A. Вид спереди, обратите внимание на широкое округлое отверстие между ушками. Асимметрия ушек и отверстия, по-видимому, связана с посмертной деформацией раковины. B — вид сбоку.
Между тем, по всей видимости, именно воронка была основным движетелем аммоноидей. У современных головоногих существует несколько механизмов плавания. Так, наутилиды в спокойном состоянии плавают за счет ундуляции воротниковых складок (так же могли плавать и аммониты), но этот способ позволяет перемещаться только с очень низкой скоростью. Если наутилусам нужно развить большую скорость, они резко втягивают голову в раковину и голова, как поршень, выжимает воду из мантийной полости через воронку, а моллюск быстро движется в противоположную сторону (Несис, 2005). Долгое время этот механизм считался универсальным для всех наружнораковинных головоногих. Но для такого поршневого движения нужны очень больше области прикрепления ретракторов головы, а это, как говорилось выше, особенность отряда Nautilida, ни у кого из цефалопод, кроме наутилид, таких областей прикрепления ректракторов головы не было. Не было их и у аммонитов и, следовательно, они не могли плавать так же, как наутилусы (Jacobs, Landman, 1993). У колеоидей механизм плавания другой - для быстрого движения они выжимают воду из мантийной полости за счет резкого сокращения мышц мускулистой мантии. Могли ли аммоноидеи иметь такой же механизм плавания? Подобные предположения высказывались (Jacobs, Landman, 1993), однако это представляется крайне маловероятным, так как мантия аммонитов явно имела не очень большой объем, помещаясь в жилой камере, и при этом ее край отвечал за строительство раковины, а совмещение таких функций вряд ли возможно (Saunders, Ward, 1994). Для медленного плавания многие современные колеоидеи используют плавники, однако у аммонитов с их наружными раковинами плавников быть не могло. Также, некоторые глубоководные осьминоги используют для передвижения в воде перепонку-мембрану, натянутую между руками, возможно, что также плавали и некоторые аммониты, однако у таких осьминогов воронка в значительной степени редуцирована (Несис, 2005), а у аммонитов она была, судя по всему, прекрасно развита.
Наличие развитой воронки у аммонитов дает основание предполагать, что именно воронка отвечала за гидрореактивное плавание этих моллюсков. То есть у аммоноидей вода могла выбрасываться из воронки не за счёт сокращения мантийной полости, а за счёт резкого сжатия самой воронки (Saunders, Ward, 1994). Надо отметить, что у колеоидей и даже у наутилусов мускулатура воронки способна в некоторой степени сжиматься и выталкивать воду (Несис, 2005). Такой механизм, конечно, был значительно менее эффективен, чем сжимание всей мантийной полости, но зато он позволял аммоноидеям реализовывать практически любую форму раковины и длину жилой камеры. Размеры и форма жилых камер, а следовательно, и раковин наутилид довольно сильно ограничены их поршневым механизмом плавания, в то время как трудно представить себе форму раковины, которую не реализовали бы те или иные аммоноидей. Возможно, что именно плавание за счет пульсаций мощной воронки позволило им быть столь разнообразными.
Голова и глаза
Полные отпечатки головы аммоноидей до сих пор неизвестны, однако, недавно в меловых отложениях Германии были найдены отпечатки аммонитов Baculites, на которых удалось рассмотреть остатки головного хряща и, возможно, глазных капсул (Klug et al., 2012; Klug and Lehmann, 2015). Судя по этим находкам, голова аммонитов была довольно крупной, примерно как у кальмаров соизмеримого размера. Хотя глазные капсулы этих образцов сохранились плохо и по ним сложно судить о строении глаз, в настоящее время исследователи не сомневаются, что аммоноидеи обладали очень хорошим зрением и имели развитые глаза (Klug et al., 2012). Это следует из того факта, что большинство из них обитали в эпипелагиали (Westermann, 1996), в хорошо освещённой солнцем воде. Кроме того, недавно было установлено, что примитивные глаза современных наутилусов — это не исходный, древнейший вариант глаз головоногих, как считали ранее, а результат деградации, упрощения глаза вследствие жизни на плохо освещённых глубинах (Ogura et al., 2013). Предки же наутилид, как и последние общие предки всех современных цефалопод, судя по всему, имели хорошо развитые глаза, не уступавшие глазам современных колеоидей.
Внутренние органы
Лучше всего из внутренних органов аммоноидей сохраняются пищевод другие элементы пищеварительной системы (Klug et al., 2012; Klug, Lehmann, 2015). У некоторых современных колеоидей стенки пищевода жесткие, укрепленные слоями хитина (Nixon, Young, 2003). Возможно, что также был устроен и пищевод аммоноидей, что и обеспечивало хорошую сохранность находок. Кроме того, хорошей сохранности пищевода и желудка способствовали остатки пищи, ускорявшие фосфатизацию тканей. Известны находки мелких фрагментов криноидей, ракообразных, раковин аммонитов и их челюстей в жилых камерах аммоноидей, интерпретированных как содержимое желудка (Tanabe et al. 2015 a).
Значительно реже и хуже сохраняются жабры аммоноидей. Известно несколько находок, интерпретированных как жабры или их фрагменты (Lehmann, 1967a, 1979, 1985, Mironenko, 2015 b), однако полной уверенности в том, что это именно жабры пока нет, да и какие-либо выводы о строении и размерах жабр по этим находкам сделать довольно сложно.
Среди других внутренних органов аммоноидей в литературе несколько раз упоминался чернильный мешок (Lehmann, 1967b). Предположения о его наличии у аммоноидей появились после того, как было установлено их близкое родство с колеоидеями, у большинства из которых чернильный мешок есть. Однако, дальнейшие исследования находок предполагаемых чернильных мешков аммоноидей показали ошибочность более ранних выводов (Lehmann, 1985; Klug, Lehmann, 2015) и сейчас нет никаких сомнений в том, что аммоноидеи не имели чернильного мешка. Иногда в качестве доказательства наличия чернильного мешка рассматривались молекулы меланина, найденные в жилых камерах аммоноидей (Doguzhaeva et al., 2004), однако, меланин входит в состав чёрного (морщинистого) слоя, покрывавшего изнутри стенки жилой камеры (о нем речь пойдет ниже) и чёрной полосы, украшавшей устье многих аммоноидей (Klug et al. 2007), поэтому его присутствие в жилой камере вовсе не свидетельствует о присутствии чернильного мешка.
Вероятнее всего, наружнораковинные головоногие моллюски в принципе не имели чернильных мешков, так как меланин, возникавших в ходе их метаболизма, отлагался на раковине в составе морщинистого слоя или в периостракуме. Только после погружения раковины под мантию у колеоидей, когда отпала необходимость в формировании морщинистого слоя и началась постепенная редукция раковины, меланин начал вводиться наружу через пищеварительную систему, что и заложило основы для формирования чернильного мешка.
Задняя часть мантии
Благодаря тому, что на раковинах аммоноидей сохраняются области прикрепления мягких тканей, исследователи смогли получить довольно много информации о задней части мантии аммоноидей. Несмотря на существенные отличия в строении мускульной систему у аммоноидей и наутилид, в задней части жилой камеры ретракторы головы у аммонитов крепились к отпечаткам мускулов примерно так же, как и у наутилид, а задняя часть мантии крепилась к раковине вдоль паллиальной линии, как и у современных наутилусов (Klug et al., 2007 b; Doguzhaeva, Mutvei, 1996). Область прикрепления задней части мантии иногда хорошо видна на ядрах раковин. Сами мягкие ткани аммонитов крайне редки в ископаемом состоянии, однако известен уникальный экземпляр Cadoceras stupachenkoi Mitta из нижнего келловея р.Унжа (Костромская область) с сохранившимся фрагментом мантийной мускулатуры и отпечатками миоадгезивной линии (Mironenko, 2015 b). Линия прикрепления задней части мантии у аммоноидей иногда посмертно обызвествляется, вероятно из-за того, что в этой части тела аммонитов содержалось много кальция, необходимого для строительства новых перегородок фрагмокона.
Морщинистый слой
Мягкое тело аммоноидей соединялось с раковиной не только в областях постоянного прикрепления мускулов-ретракторов и паллиальной линии. Мантия аммоноидей могла прикрепляться к внутренним стенкам жилой камеры и поверхности предыдущего оборота за счет специального морщинистого слоя (wrinkle layer), располагавшегося поверх гладкого арагонита раковины. Этот слой имел морщинистую шероховатую поверхность и позволял мантии крепко держаться за раковину в покрытых им областях (Догужаева, 1981; Keupp, 2000; Klug et al., 2007b). У современных наутилусов гомологом морщинистого слоя является, по-видимому, органический чёрный слой, также имеющий морщинистую поверхность и покрывающий небольшой участок предыдущего оборота вне жилой камеры, перед устьем, а также устьевой край у взрослых особей (Ward, Saunders, 1997, фиг. 1). У аммонитов же морщинистый слой покрывал не только узкую полосу вдоль устья, но и всю внутреннюю поверхность жилой камеры (кроме областей постоянного прикрепления мускулатуры) и точно так же, как и у наутилусов, захватывал участок предыдущего оборота вне устья (Klug et al., 2007b; Mironenko, 2015c).
Строение морщинистого слоя, рельефность и форма его поверхности у разных таксонов аммоноидей существенно различались. Так, у некоторых аммоноидей этот слой полностью состоял из органического вещества (как и у современных наутилусов) и имел лишь слегка шероховатую поверхность, в то время как у других он включал арагонитовые вкрапления и был покрыт грубыми извилистыми рёбрами и складками (Kulicki, 1979). Причины такого разнообразия морщинистого слоя заключались, по-видимому, в различии в образе жизни аммоноидей, обладающих морщинистым слоем разных типов. Особенно сильно различался участок морщинистого слоя, расположенный вне жилой камеры, на поверхности предыдущего оборота раковины. Этот участок, названный «затылочной прикрепительной областью» (supracephalic attachment area – см. Mironenko, 2015c) был обнаружен на раковинах многих аммонитов (Догужаева, 1981; Birkelund, 1981; Keupp, 2000; Klug et al., 2007b), однако до недавнего времени это были единичные находки и палеонтологи не придавали им практически никакого значения. Комплексное исследование этих площадок стало возможно благодаря находкам раковин аммонитов хорошей сохранности с морщинистым слоем в средне-верхнекелловейских и верхнеоксфордских отложениях Центральной России: в Михайлове (Рязанская область), Дубках (Саратовская область) и в Марково (Московская область) (Рис.4).

Рис. 4. Сохранившаяся затылочная прикрепительная область на раковине Quenstedtoceras lamberti (показана фигурной скобкой), средняя юра, верхний келловей, Саратовская область, Дубки. А — общий вид сохранившегося образца, B — реконструкция, показывающая не сохранившуюся жилую камеру и положение прикрепительной области относительно устья. По Mironenko, 2015 c с изменениями.
Было установлено, что характер рельефа морщинистого слоя и размер затылочной прикрепительной площадки напрямую связаны с формой раковины (Mironenko, 2015c). Так, образованные морщинистым слоем прикрепительные площадки на оксиконических и дискоконических инволютных раковинах отличаются значительно большей площадью и более грубым рельефом по сравнению с площадками на платиконических эволютных раковинах. В качестве примера можно привести гладкие обтекаемые раковины келловейского рода Sublunuloceras и ребристые, украшенные многочисленными шипами раковины Kosmoceras. У Kosmoceras длина прикрепительной площадки составляет 20-22 градуса окружности, а рельеф морщинистого слоя виден только под большим увеличением, в то время как на раковине Sublunuloceras такого же размера с такой же длиной жилой камеры длина площадки превышает 70 градусов окружности, а ее поверхность покрыта сетчатым орнаментом, хорошо заметным невооружённым глазом (Mironenko, 2015c).
Форма раковины аммонитов связана с их образом жизни и в первую очередь со скоростью и маневренностью плавания (Jacobs, Chamberlain, 1996). Обтекаемые инволютные раковины были лучше приспособлены к маневрированию и быстрому ускорению, чем эволютные и особенно покрытые грубыми ребрами и шипами формы. Зависимость между формой и рельефом затылочной площадки с одной стороны и формой раковины с другой говорит о том, что именно для аммонитов с обтекаемыми раковинами важно было иметь наиболее сильное сцепление мягких тканей с поверхностью раковины и это сцепление как раз обеспечивалось затылочной прикрепительной областью.
Воротник
Наличие затылочной прикрепительной области вне жилой камеры аммоноидей также проливает свет на строение мягкого тела этих моллюсков. У современных наутилусов чёрный слой перед устьем выделяется затылочной складкой мантии (Klug et al., 2007). Наличие прикрепительной площадки, сформированной морщинистым слоем на раковинах аммоноидей свидетельствует о том, что у них тоже была затылочная складка мантии. Однако, этот участок мантии не подвергается особым нагрузкам и для его фиксации на поверхности раковины не нужно такое рельефное строение прикрепительной области, какое наблюдается у многих юрских аммонитов. Высказывались предположения, что рельефные площадки были нужны для прикрепления капюшона (Klug et al., 2004), однако, капюшона, как у наутилусов, у аммоноидей быть не могло, это образование характерно только для наутилид (подробнее см. Mironenko, 2015c). В состав капюшона наутилид кроме двух пар щупалец и тканей глаза входит еще одна очень важная структура — дорсальная часть воротника (Shigeno et al., 2008, 2010).
Воротник цефалопод крайне редко упоминается в палеонтологической литературе, однако, это важнейшая часть мягкого тела головоногих моллюсков (Bizikov, 2002, Shigeno et al., 2008, 2010). По-видимому, именно ткани воротника отвечают у наутилуса за сцепление капюшона с чёрным слоем. Воротниковые складки, расположенные по бокам позади головы, у всех головоногих отвечают за дыхание и поступление воды в мантийную полость, в которой, среди прочего, располагаются и жабры моллюска (Bizikov, 2002, 2008). На вентральной стороне воротниковые складки переходят в воронку (Bizikov, 2002, 2008), то есть воронка — это не отдельная трубка, вставленная в мантийную полость, как ее иногда рисуют, а по сути вырост воротника. На дорсальной стороне воротниковые складки срастаются между собой и у колеоидей соединяются с головой, а у наутилид входят в состав капюшона. Однако, на эмбриональной стадии развития у всех головоногих моллюсков воротник обособлен (Shigeno et al., 2008, 2010). У аммоноидей, не имевших капюшона, он, скорее всего, должен был оставаться обособленным на протяжении всего онтогенеза (хотя его дорсальные края могли срастаться с головой). По всей видимости, именно воротник прикреплялся к затылочной прикрепительной области. Это объясняет обнаруженную взаимосвязь между формой раковины (и как следствие активностью животных) и размерами затылочной прикрепительной площадки. Воротник был непосредственно связан с воронкой и все нагрузки, возникавшие при выбрасывании воды из воронки, тут же передавались на ту часть воротника, которая «держалась» за прикрепительную площадку. То есть аммонит не высовывался из жилой камеры как червяк из норы (что часто можно увидеть на палеореконструкциях), а крепко охватывал часть предпоследнего оборота своей раковины перед устьем (реконструкция внешнего вида аммонитов с учетом прикрепительной области — Mironenko, 2015c, фиг. 18). Такая связь позволяла аммоноидеям обеспечивать надежное управление движением: при ускорении или маневрировании импульс мгновенно передавался на раковину. А в случае опасности аммонит, как и современный наутилус, мог практически мгновенно отцепиться от прикрепительной области и скрыться в жилой камере.
Прижизненная ориентация раковин в воде
Крупные размеры прикрепительных площадок, достигающие 100 градусов окружности (у микроконхов Pseudocadoceras) позволяют уточнить еще один важный аспект палеобиологии аммонитов — прижизненную ориентацию их раковин. Ориентация любого объекта в воде определяется соотношением центров тяжести и плавучести. В 1941 году Трумэн (Trueman, 1941) предложил методику расчета положения аммонитов в воде, которая с некоторыми незначительными уточнениями и дополнениями применяется до сих пор (Tajika et al., 2015). В основе предложенных Трумэном расчетов лежало допущение, что мягкое тело аммонитов было однородной белковой массой, занимавший весь объём жилой камеры. Ни тот факт, что внутри тела аммонитов находилась заполненная водой мантийная полость, ни способность аммонитов высовываться из раковины в расчёт не принимались, поскольку влияние этих факторов было крайне трудно оценить (Trueman, 1941). Однако, результаты таких расчётов для многих аммонитов оказались весьма странными: получилось, что их устьевой край будто бы был расположен горизонтально, отверстие устья смотрело вверх, на поверхность воды. В такой позе и воронка, и щупальца оказывались бы направлены вертикально вверх (это можно увидеть на некоторых реконструкциях: Westermann, 1996, фиг 5; De Baets et al., 2013, табл. 1). Особенно странно это выглядело для аммонитов с вентральным выступом-ростром на устье, так как он тоже должен был бы торчать вертикально вверх, мешая воронке хотя бы перегнуться через край раковины. Некоторые авторы, включая самого Трумэна, отмечали несовершенство используемой методики, но так как объём мантийной полости и той части тела, которая в активном состоянии выдвигалась из жилой камеры было очень сложно оценить, все продолжали вести расчеты по-старому, внося незначительные уточнения (учитывая, к примеру, вес тканей сифона или шипов на раковине). Однако, сформированные морщинистым слоем прикрепительные площадки позволяют нам уточнить, какая часть мягкого тела (как минимум) выдвигалась за пределы устья. Так как к прикрепительной площадке присоединялся воротник, который всегда расположен позади головы, нет сомнений, что вся голова вместе с руками, челюстным аппаратом и воронкой выдвигались из жилой камеры наружу. Соответственно, можно попробовать оценить и объём мантийной полости, который не должен был быть меньше объёма выдвигавшихся наружу мягких тканей. Такие расчеты еще не были проведены, но это дело ближайшего будущего.
Аптихи
Еще одним фактором, который может помочь уточнить объём тела аммонитов и размер отдельных его частей, является размер аптихов. Согласно современным представлениям, аптихи — это нижние челюсти аммонитов, которые при этом могли использоваться и как крышечки для защиты просвета жилой камеры (Lehmann, Kulicki, 1990; Parent, Westermann, 2016). Первоначально под названием «аптихи» были описаны парные кальцитовые пластинки, расположенные на внешней поверхности аммонитовых челюстей аптихового типа, однако чаще всего в литературе аптихами называют нижние челюсти (или их половинки) целиком, даже если они не имели кальцитового слоя на поверхности.
Параллельно с аммонитами, обладавшими челюстным аппаратом аптихового типа, в юре у аммоноидей продолжали существовать и другие типы челюстных аппаратов — анаптиховый и ринхаптиховый. Подробный обзор и сравнение различных типов челюстных аппаратов можно найти в публикации К.Танабе с соавторами (Tanabe et al. 2015a).

Рис. 5. Аптих аммонита Mirosphinctes, верхняя юра, верхний оксфорд, Московская область, Рыбаки. Сохранился только органический слой аптихи, кальцитовый слой на данном образце разрушен. А. - общий вид аптиха, B — прижизненное повреждение (похожее на небольшой «шрам») на органической части аптиха.
До недавнего времени аптихи аммонитов с территории Центральной России в целом и Московской области в частности известны не были, однако, в течение последних полутора десятков лет ситуация существенно изменилась. Аптихи были найдены в келловейских отложениях в карьере Михайловцемент в Рязанской области (Рогов, 2004 a), в оксфордских отложениях у д. Рыбаки в Московской области (Rogov, Mironenko, 2016; Mironenko, 2017a, Рис. 5) и в волжских отложениях в Среднем Поволжье (Рогов, 2004b), а также в карьере Еганово в Московской области и Карамышевская набережная на территории Москвы (Мироненко, 2014). Оксфордские и волжские аптихи были обнаружены в жилых камерах аммонитов, которым они принадлежали, что позволило сравнить размер аптихов с диаметром устья аммонитов. Оказалось, что у волжских Kachpurites и оксфордских микроконхов Mirosphinctes длина и ширина аптихов практически совпадают с длиной и шириной устьевого отверстия жилой камеры, а вот у оксфордских макроконхов Euaspidoceras аптихи примерно на 20% меньше устья (Rogov, Mironenko, 2016). Подобная картина была обнаружена и у других аммонитов (Keupp, Mitta, 2013; Larson, Landman, 2017), в некоторых случаях длина аптихов даже на 30% меньше высоты устья (Larson, Landman, 2017). Хотя нельзя исключать того, что при достижении определенного размера аммонита, необходимость защитной функции аптихов исчезала (большинство аммонитов с аптихами, которые меньше устья — макроконхи), размер аптихов можно попытаться использовать для оценки объёма той части жилой камеры, которую могло занимать тело аммонита позади аптихов. Изучение залеченных повреждений на раковинах аммонитов показало, что многие аммониты были способны пережить разрушение почти половины жилой камеры начиная от устья, причем без видимых травм мантийного края (Kröger, 2002). Это свидетельствует о способности аммонитов очень глубоко втягиваться в жилую камеру (Kröger, 2002) и является дополнительным подтверждением того, что объём их мягкого тела вовсе не равнялся объёму жилой камеры. Кроме того, многие современных гастроподы, имеющие крышечку, перекрывают этой крышечкой просвет раковины на некотором расстоянии от устья (Kaim, Sztajner, 2005).
Размеры аптихов позволяют не только уточнить объём жилой камеры, который во втянутом в нее состоянии занимало мягкое тело аммонита, но и длину головы моллюска. Поскольку аптихи являлись частью челюстного аппарата, то голова аммонита не могла быть короче аптихов, это можно учитывать при реконструкции внешнего вида аммонитов.
Аптихи сочетали в себе как минимум две функции: защитную и челюстную (возможно, были и другие, см. Parent, Westermann, 2016). Необходимость этого совмещения накладывала на них свой отпечаток и в качестве нижней челюсти они функционировали, скорее всего, не столь эффективно, как нижние челюсти аммоноидей, относящиеся к другим челюстным типам (анаптиховому и ринхаптиховому). Однако, челюстной аппарат аммоноидей включал не только челюсти (верхнюю и нижнюю), но и радулу: расположенную между челюстями ленту, покрытую множеством мелких зубчиков (Keupp et al., 2016). Зубчики расположены рядами, у аммоноидей, как и у большинства колеоидей, в каждом ряду было по 7 зубчиков (у наутилид их 9). Недавно было установлено, что по крайней мере у некоторых аммонитов, обладавших аптихами, форма этих зубчиков отличалась от таковой у всех остальных аммоноидей: зубчики таких аммонитов имели значительно более сложное строение и обладали множеством отдельных вершин. Вполне вероятно, что именно значительное усложнение радулы несколько «разгрузило» нижнюю челюсть и позволило аммонитам использовать её для защиты жилой камеры, пусть и с некоторым ущербом для основной функции как части челюстного аппарата (Keupp et al., 2016).
Щупальца
Конечно, важнейшей частью головного отдела всех головоногих моллюсков является их ручной аппарат. Именно его форма и строение лучше всего свидетельствуют об образе жизни моллюска. К сожалению, до сих пор не найдено ни одного отпечатка ручного аппарата аммоноидей: их рук или щупалец. Точнее, хотя такие находки несколько раз упоминались в литературе (De la Torre y Capablanca 1953; Stürmer, 1969; Zeiss, 1969), но сейчас исследователи полагают, что в этих случаях за отпечатки рук аммоноидей были приняты либо ихнофоссилии — ходы и следы ползания животных, живших в заполнившем раковину осадке, либо неоднородности самого осадка (Klug, Lehmann, 2015).
Однако, благодаря изучению эмбрионального развития современных головоногих, некоторые выводы о ручном аппарате аммоноидей сделать можно. Изучение эмбрионов всех современных цефалопод, включая наутилусов, показывает, что исходное число щупалец у них всегда равно десяти (Shigeno et al., 2008, 2010). У наутилусов эти десять исходных зачатков щупалец позже многократно дробятся, но такое строение ручного аппарата возникло у них уже после того, как разошлись эволюционные ветви наутилоидей и других головоногих. Cейчас мы с большой уверенностью можем утверждать, что изначально аммоноидеи, также как и их общие с колеоидеями предки — бактритиды, имели 10 рук (Klug, Lehmann, 2015). Что было дальше, сохранили ли все аммоноидеи это исходное число рук на протяжении всей их эволюции или оно изменялось — это пока остается открытым вопросом. Может быть у кого-то из аммоноидей, как и у осьминогов, одна пара конечностей редуцировалась, а может быть наоборот, удвоилось число рук, но пока не будут обнаружены достоверные отпечатки, большинство исследователей пока сходится на том, чтобы считать число рук аммоноидей равным десяти (Рис. 6).
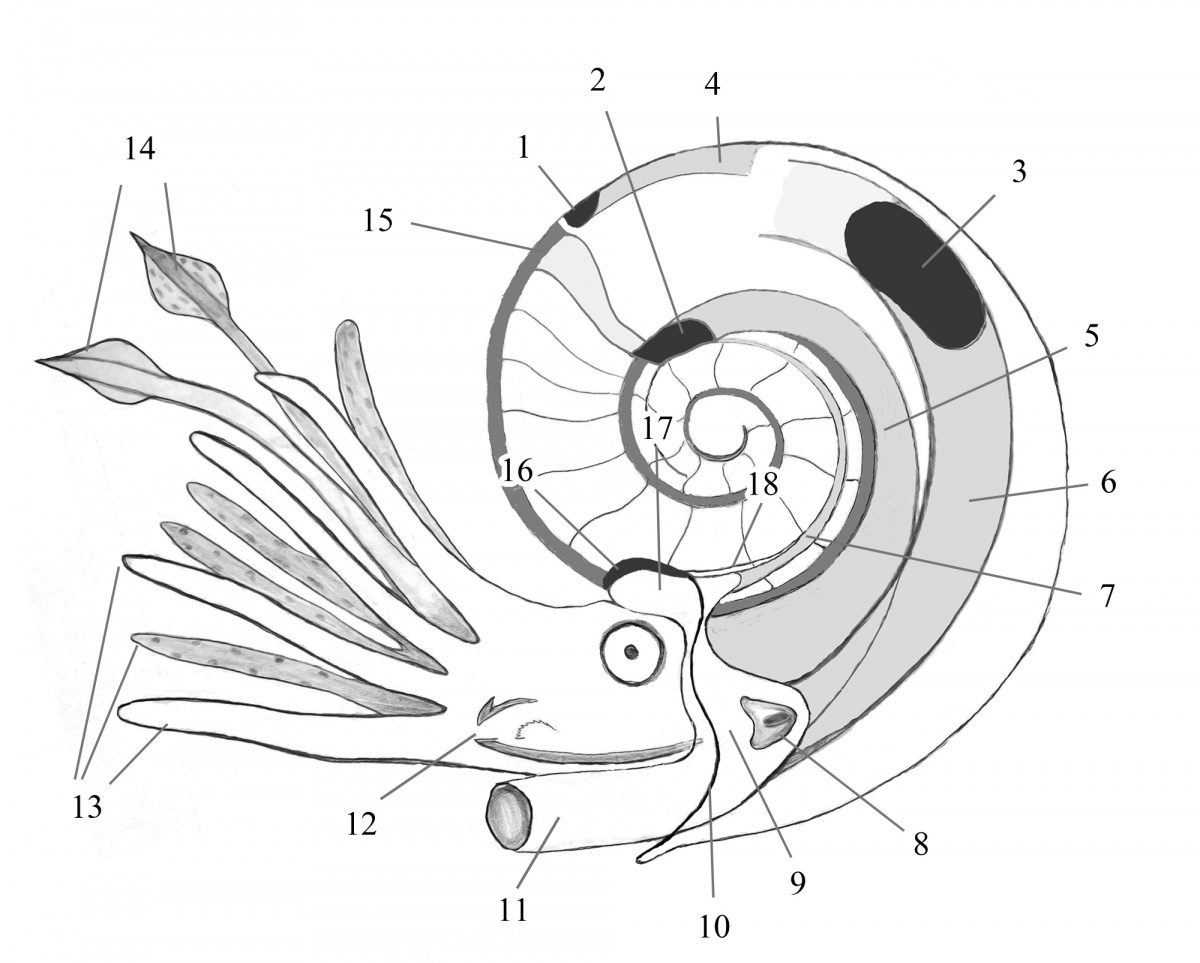
Рис. 6. Схематическая реконструкция мускульной системы и мягких тканей аммонитов. На основе схемы Mironenko 2015 a, фиг.13 (с изменениями). Жилая камера и фрагмокон показаны в разрезе, но показана затылочная прикрепительная область (морщинистый слой), расположенная на наружной поверхности раковины около устья. Цифрами обозначены: 1 - вентральный отпечаток мускула, 2 - дорсо-латеральный отпечаток мускула, 3 - латеральный отпечаток мускула, 4 - вентральный мускул, 5 - ретрактор головы, 6 - ретрактор воронки, 7 - затылочный ретрактор, 8 - мантийно-вороночный замыкатель, 9 - воротник, 10 - устье раковины, 11 - воронка, 12 - челюсти и радула, 13 - руки, 14 - щупальца, 15 - сифон, 16 - затылочная прикрепительная область (морщинистый слой), 17 - затылочные участки мантии и воротника, 18 - пупковый шов.
Еще один вопрос, касающийся ручного аппарата аммоноидей: были ли руки аммоноидей одинаковыми, или две из них могли преобразоваться в хватательные щупальца, как у каракатиц и некоторых кальмаров? Наличие хватательных щупалец существенно облегчает охоту головоногих моллюсков, особенно при атаке на добычу, находящуюся впереди, поскольку наибольшую скорость головоногие развивают при движении назад, быстро двигаться головой вперед им сложнее. Аммоноидеям с их наружной раковиной бросаться на добычу головой вперед наверняка было еще труднее, чем каракатицам с их обтекаемым телом, и появление хватательных щупалец у аммоноидей представляется весьма логичным и вероятным, поэтому их часто изображают на реконструкциях (Mironenko, 2015c, фиг. 18).
Многие исследователи предполагают, что аммониты с аптиховым типом челюстного аппарата питались в основном некрупной добычей и планктоном (Kruta et al., 2011). Хотя такой образ жизни вряд ли стоит экстраполировать на всех аммонитов с аптиховыми челюстями, он, тем не менее, представляется вполне вероятным для большинства гетероморфных аммонитов и аммонитов с широкими и скульптурированными раковинами, например для многих перисфинтид. Современные головоногие, питающиеся планктоном и мелкоразмерной добычей имеют специальную тонкую мембрану между руками, помогающую удерживать частицы пищи и собирать их. У некоторых колеоидей такая мембрана доходит до середины рук, у других — почти до самых кончиков (Несис, 2005). Наличие такой мембраны у аммонитов-планктонофагов выглядит логичным и вполне вероятным (Рис. 7).

Рис. 7. Реконструкция внешнего вида микроконха Indosphinctes sp., средняя юра, средний келловей, Центральная Россия. Показаны хорошо развитые глаза, мощная воронка и десять рук, соединенных тонкой мембраной. На реконструкции показана гипотетическая яркая окраска боковых выростов устья (ушек), которая подчеркивает зрелость аммонита и его готовность к размножению (предполагается, что микроконхи — раковины самцов аммонитов, в то время как макроконхи — раковины самок). Рисунок Андрея Атучина по эскизу автора статьи.
Кровеносные сосуды сифона
Говоря об анатомии и палеобиологии аммоноидей, стоит упомянуть и те мягкие ткани, которые располагались не в жилой камере, а во фрагмоконе. В первую очередь, это кровеносные сосуды сифона. До недавнего времени были известны только две находки фосфатизированных структур внутри сифонных трубок, которые были интерпретированы как остатки кровеносных сосудов — в раковинах Akmilleria electraensis из перми США (Tanabe et al., 2000, 2015b) и Virgatitus virgatus из волжского яруса верхней юры, найденного на территории Москвы (Барсков, 1990). Однако, строение структур, обнаруженных внутри сифонных трубок, в этих двух случаях оказалось различным: у Akmilleria число и расположение сосудов были сходны с числом и расположением сосудов в сифоне современного Nautilus (Tanabe et al., 2000, 2015b), а у Virgatites количество трубок, интерпретированных как остатки сосудов, менялось на разных оборотах раковины и не совпадало с числом сосудов сифона у современных цефалопод (Барсков, 1990). Недавно удалось обнаружить фосфатизированные фрагменты мягких тканей сифонов еще у одного вида аммонитов — верхневолжского Kachpurites fulgens, причем остатки сосудов были найдены сразу у нескольких экземпляров (Mironenko, 2017b). Хотя форма сосудов у Kachpurites оказалась довольно сильно искажена отложениями апатита, их число и расположение отчётливо видны и совпадают с числом и расположением сосудов в сифоне современного наутилуса и пермской Akmilleria (Mironenko, 2017b). Это подтверждает высказанное ранее мнение о консервативности строения сифонных сосудов у аммонитов и о том, что в сифоне Virgatites, к сожалению, были найдены не сосуды, а вторичные фосфатные образования (Tanabe et al., 2000, 2015b).
Интересно отметить, что несмотря на сходство в числе и расположении кровеносных сосудов, мягкие ткани сифонов аммоноидей (и у Kachpurites и у Akmilleria) отличаются от мягких тканей сифонов наутилусов числом складок наружного эпителия (Mironenko, 2017 b). Число борозд и гребней на эпителии соответствует количеству особых клеток, отвечающих за газо-жидкостный обмен между сифоном и камерами фрагмокона, чем их больше, тем быстрее должен работать сифон (Tanabe et al., 2000, 2015b). У современного наутилуса число таких борозд и гребней почти на порядок больше, чем у аммоноидей. Но при этом, существуют многочисленные свидетельства того, что аммоноидеи должны были уметь быстро менять плавучесть своих раковин, к примеру, для того, чтобы скомпенсировать потерю части раковины после травмы (Kröger, 2002). Значит, несмотря на меньшее число необходимых для работы сифона клеток, аммоноидеи либо имели какие-то дополнительные механизмы, обеспечивающие улучшение работы сифона, либо существенную роль играли мембраны, которые присутствовали во фрагмоконах аммоноидей, но отсутствуют у наутилид.
Мембраны
Мембраны, которые могли способствовать улучшению работы сифона аммоноидей — загадочные образования, функции и происхождение которых еще не до конца ясны (Polizzotto et al., 2015). Однако известно, что эти органические мембраны делили каждую камеру фрагмокона на небольшие отсеки, расположенные вокруг сифона. Подобное расположение мембран должно было удерживать жидкость в камерах фрагмокона в непосредственной близости от сифонной трубки, что наверняка позволяло быстрее откачивать её из камер, чем если бы жидкость могла растекаться по всему объёму камеры. Хорошая сохранность мембран даже в тех камерах, где разрушена значительно более толстая сифонная трубка наводит на мысль о том, что мембраны были, несмотря на малую толщину, очень плотными и состояли из весьма устойчивого органического вещества. А значит, основной их функцией была не транспортировка воды к сифону, как это иногда предполагалось, а разделение камер на отдельные отсеки. Мембраны также обеспечивали прикрепление сифона к стенке раковины. Они хорошо сохраняются в раковинах волжских и келловейских аммонитов, найденных в Московской, Ярославской и Костромской областях. Особенно хорошо мембраны видны на кальцитовых ядрах раковин аммонитов Craspedites из местонахождения Огарково в Костромской области (Рис. 8).

Рис. 8. Края мембран (тонкие чёрные линии), видимые между перегородками фрагмокона на ядре аммонита Craspedites, верхняя юра, верхняя волга, Огарково, Костромская область. Форма мембран несколько различается в каждой камере, видимо, из-за деформации органических мембран в ходе разложения и фосфатизации.
Размножение аммоноидей
В последние годы удалось уточнить не только анатомические особенности аммоноидей, но и некоторые аспекты их образа жизни. Так, было показано, что аммоноидеи (по крайней мере юрские и меловые аммониты) не откладывали яйца, а были живородящими. Яйцеживорождение известно у двух монотипических родов современных пелагических осьминогов Ocythoe и Vitreledonella (Несис, 2005), причем эти таксоны не являются близкими родственниками и такой путь размножения у них, по-видимому, возник независимо друг от друга. Ocythoe относятся к надсемейству Argonautoidea, два других представителя этого надсемейства — Argonauta и Tremoctopus также не откладывают яйца в воду или в какие-либо укрытия, а постоянно носят их с собой до вылупления потомства (самки Tremoctopus переносят их в руках, а самки Argonauta — в специальной раковине). При этом и у живородящих Ocythoe, и у Argonauta с Tremoctopus развит экстремальный половой диморфизм, при котором самки в 2-10 раз (Несис, 2005), а у Tremoctopus в некоторых случаях и почти в 100 раз (Norman et al., 2002) превосходят по размеру самцов Неоднократно высказывались предположения о том, что аммониты также могли быть живородящими, то есть яйца развивались в жилых камерах самок до выхода молоди. В качестве свидетельств в пользу этой гипотезы рассматривались и половой диморфизм юрских и меловых аммонитов, очень сходный с диморфизмом современных аргонаутоидей, и многочисленные находки сферических объектов в жилых камерах, которые интерпретировались как остатки икры, вынашивавшейся внутри тела аммоноидей (Баранов, 1985; Landman et al., 2010). Правда, одна из таких находок (Баранов, 1985) оказалась яйцевыми капсулами гастропод, отложенными в жилую камеру аммонита уже после смерти моллюска (Zatoń, Mironenko, 2015), да и в остальных случаях оставались сомнения в природе этих находок.
Но недавно в отложениях аптского яруса нижнего мела Саратовской области (у д. Широкий Буерак) были найдены раковины аммонитов Sinzovia с многочисленными эмбриональными раковинами (аммонителлами) в жилых камерах (Mironenko, Rogov, 2016). Сохранившиеся кальцитовые нижние челюсти аммонителл и отсутствие каких-либо иных фрагментов или раковин в этих скоплениях свидетельствовали о том, что это действительно остатки эмбрионов, находившихся в жилых камерах аммонитов в момент их гибели, то есть эти аммониты были живородящими (Mironenko, Rogov, 2016). Раковины были найдены в чёрных сланцах, формировавшихся в бескислородных условиях, по-видимому аммониты с аммонителлами в жилой камере погибли во время подъема бескислородных вод к поверхности.
Вопрос о времени возникновения яйцеживорождения у аммоноидей пока остается открытым. В самом конце девона эмбриональные раковины аммонитов приобрели форму и размер (1-2 мм), остававшиеся неизменными на протяжении всей их дальнейшей эволюции, и возможно, что это было связано с появлением живорождения. Но с другой стороны, заметный диморфизм, который также мог быть следствием возникновения живорождения, появился только в конце ранней юры. Возможно, что будущие исследования помогут ответить на вопрос о появлении живорождения у аммоноидей.
Заключение
Несмотря на множество открытий в области палеобиологии и анатомии аммоноидей, здесь все еще остается много нерешенных вопросов. Мало известно о строении верхней челюсти в аптиховом и ринхаптиховом типах челюстных аппаратов аммоноидей, а без этой информации сложно объективно судить об их пищевом поведении. До сих пор не найдено ни одного отпечатка рук или воронки аммоноидей. Хотя наружная раковина и препятствовала сохранению мягкого тела, такие находки вполне возможны, например, если раковина была разрушена в результате атаки хищника, а мягкое тело частично уцелело и было захоронено в донном осадке. Для поиска таких образцов особое внимание стоит обратить на чёрные сланцы, в которых встречаются остатки мягких тканей других животных, таких как колеоидеи или рыбы. Мало известно о причинах возникновения различных аномалий раковин аммоноидей, не связанных с травмами или с видимым воздействием паразитов или обрастателей (асимметрия фрагмокона, асимметрия и инверсия лопастной линии). Есть и другие нерешенные вопросы. Но интенсивность исследований аммоноидей и обилие хороших находок позволяют надеяться, что в ближайшие годы многие из вопросов будут разрешены и мы узнаем много нового о жизни этих замечательных древних животных.
Благодарности
Автор благодарен Ирине Смуровой (Москва) за изготовление фотографий к этой публикации, Омару Хаписову (Дагестан) за переданные для исследования образцы и Михаилу Рогову (ГИН РАН) за ценные замечания и дополнения. Также автор благодарит палеоиллюстратора Андрея Атучина за создание реконструкции внешнего облика аммонита. Работа выполнена по теме ГИН РАН № 0135-2014-0064 при поддержке гранта РФФИ № 05-15-06183.
Литература
Баранов В.Н. Об остатках икры в жилых камерах поздневолжских аммонитов // Бюллютень МОИП. Отдел геологический. 1985. Т. 60. №2. С. 89-91.
Барсков И.С. Внутреннее строение сифона позднеюрского аммонита Virgatites virgatus (Buch) // Труды Палеонтологического института. 1990. Т. 243. С. 127-135.
Догужаева Л.А. Морщинистый слой раковины аммоноидей // Палеонтологический журнал. 1981. 1. c. 38-48
Догужаева Л.А., Кабанов Г.К. Мускульные отпечатки аммоноидей // Доклады Академии наук СССР. 1988. Т.301. №1. С.210-213.
Мироненко А.А. Первая находка челюстного аппарата верхневолжских аммонитов Kachpurites fulgens (Craspeditidae) // Палеонтологический журнал,.2014. №6. С. 12-17.
Несис К.Н. Головоногие: умные и стремительные (Истории из частной и семейной жизни кальмаров, каракатиц, осьминогов, а также наутилуса помпилиуса). М.: Издательство «Октопус”. 2005. 204 с.
Рогов М.А. Средне- и позднекелловейские аптихи (Ammonoidea) Русской плиты // Палеонтологический журнал. 2004 a. №1. С. 26-33.
Рогов М.А. Аптихи из волжского яруса русской платформы // Палеонтологический журнал. 2004 b. №2. С. 28-34.
Barskov I.S. Phosphatized blood vessels in the siphuncle of Jurassic ammonites // Bulletin de L’Institut oceanographique, Monaco. 1996. Vol. 14. P. 335–341.
Birkelund T. Ammonoid shell structure // In House, M.R. & Senior, J.R. (eds.) The Ammonoidea. The Systematic Association, Special Volume 18. 1981. P. 177–214.
Bizikov V.A. Reanalysis of functional design of Nautilus locomotory and respiratory systems // American Malacological Bulletin. 2002. Vol. 17. P. 17–30.
Bizikov V.A. Evolution of the Shell in Cephalopoda. Moscow: VNIRO Publishing, 2008. 444 pp.
De Baets K., Klug C., Korn D., et al. Emsian Ammonoidea and the age of the Hunsrück Slate (Rhenish Mountains, Western Germany). // Palaeontographica. Abt. A 2013. Bd.299. P.1–114
De la Torre y Capablanca C. Dos Casos de impresiones de las partes blandas de dos Ammonoideos des Oxfordiense superior de Viñales (Cuba) // Estudios geológicos. 1953. Vol.19. P.407-414.
Doguzhaeva L.A., Mapes R. The Body Chamber Length Variations and Muscle and Mantle Attachments in Ammonoids // // In Klug C., Korn D., De Baets K., Kruta I., Mapes R.H. (eds.) Ammonoid paleobiology: from anatomy to ecology. Dordrecht: Springer, 2015. P. 545-584
Doguzhaeva L.A. Mutvei H. Organization of the soft body in Aconeceras (Ammonitina), interpreted on the basis of shell morphology and muscle scars // Palaeontographica. Abt. A. 1991. Bd. 218. P.17–33.
Doguzhaeva L.A., Mutvei H. Shell ultrastructure, muscle scars, and buccal apparatus in ammonoids. // Geobios, Memoire Special. 1993. T.26. P.111–119.
Doguzhaeva L.A., Mutvei, H. Attachment of the body to the shell in ammonoids // In: Landman, N.H., Tanabe, K., Davis, R.A. (eds.) Ammonoid paleobiology. New York: Plenum Press, 1996. P. 44–64.
Doguzhaeva L.A., Mutvei H., Summesberger H., Dunca E. Bituminous soft body tissues in the body chamber of the Late Triassic ceratitid Austrotrachyceras from the Austrian Alps // 2004. Hft. 88. P.37–50
Jacobs D.K., Chamberlain J.A. Buoyancy and hydrodynamics in ammonoids // In Landman, N., Tanabe, K., Davis, R.A. (eds.) Ammonoid Paleobiology. Topics in Geobiology 13. New York: Plenum Press. 1996. P. 169–223.
Jacobs, D.K., Landman, N.H. Nautilus — a poor model for the function and behavior of ammonoids? // Lethaia. 1993.Vol.26. no.2. P. 101-111.
Kaim A., Sztajner P. The opercula of neritopsid gastropods and their phylogenetic importance // Journal of Molluscan Studies. 2005. 71. P. 211– 219.
Kennedy W.J., Cobban W.A. Aspects of ammonite biology, biogeography, and biostratigraphy // Special Papers in Palaeontology. Vol. 17. 1976. 94 p.
Keupp H. Ammoniten. Paläobiologische Erfolgsspiralen. Stuttgart: Thorbecke Species, 2000. 165 S.
Keupp H., Hoffmann R., Stevens K., Albersdörfer R. Key innovations in Mesozoic ammonoids: the multicuspidate radula and the calcified aptychus // Palaeontology. 2016. Vol. 59. P.775-791.
Keupp H., Mitta V.V. Cephalopod jaws from the Middle Jurassic of Central Russia // Neues Jahrbuch fur Geologie und Palaontologie. Abhandlungen. 2013. Bd. 270. P.23–54.
Klug C., Korn D., Richter U., Ulrich M. The black layer in cephalopods from the German Muschelkalk (Triassic) // Palaeontology. 2004. Vol. 47. P.1407-1425.
Klug C., Lehmann J. Soft part anatomy of ammonoids: reconstructing the animal based on exceptionally preserved specimens and actualistic comparisons // // In Klug C., Korn D., De Baets K., Kruta I., Mapes R.H. (eds.) Ammonoid paleobiology: from anatomy to ecology. Dordrecht: Springer, 2015. P. 507-529
Klug C., Brühwiler T., Korn D., Schweigert G., Brayard A., Tilsley J. Ammonoid shell structures of primary organic composition // Palaeontology 2007 a. Vol.50. P.1463–1478
Klug C., Montenari M., Schulz H., Ulrich, M.. Soft-tissue attachment of Middle Triassic Ceratitida from Germany // In Cephalopods Present and Past: New Insights and Fresh Perspectives. Springer, 2007 b. P. 205-220.
Klug C., Riegraf W., Lehmann J. Soft-part preservation in heteromorph ammonites from the Cenomanian-Turonian Boundary Event (OAE 2) in the Teutoburger Wald (Germany) // Palaeontology. 2012. Vol.55. P.1307–1331.
Kröger B. On the ability of withdrawing of some Jurassic ammonoids // Abhandlungen der Geologischen Bundesanstalt. 2002. Bd. 57. P.199–204.
Kruta I., Landman N., Rouget I., Cecca F., Tafforeau P. The role of ammonites in the Mesozoic marine food web revealed by jaw preservation // Science. 2011. Vol.331. no.6013. P.70-72.
Kulicki C. The ammonite shell: its structure, development and biological significance // Acta Palaeontologica Polonica. 1979. Vol. 39. P.97–142.
Landman N.H., Mapes R.H., Cruz C. Jaws and soft tissues in ammonoids from the Lower Carboniferous (Upper Mississippian) Bear Gulch Beds, Montana, USA // In: Tanabe K., Shigeta Y., Sasaki T. & Hirano H. (Eds):Cephalopods: Present and Past. Kanagawa: Tokai University Press, 2010. 147–154.
Larson N.L., Landman N.H. Description of the lower jaws of Baculites from the Upper Cretaceous US Western Interior // Acta Geologica Polonica. 2017. Vol. 67. no.1. P.109-120.
Lehmann U. Ammoniten mit Kieferapparat und Radula aus Lias-Geschieben // Paläontologische Zeitschrift. 1967 a. Bd.41. S.38–45.
Lehmann U. Ammoniten mit Tintenbeutel // Paläontologische Zeitschrift. 1967 b. Bd. 41. S.132–136
Lehmann U. The jaws and radula of the Jurassic ammonite Dactylioceras // Palaeontology. 1979. Vol.22. P.265–271.
Lehmann U. Zur Anatomie der Ammoniten: Tintenbeutel, Kiemen, Augen // Paläontologische Zeitschrift. 1985. Bd. 59. S.99–108.
Lehmann U., Kulicki C. Double function of aptychi (Ammonoidea) as jaw elements and opercula // Lethaia. 1990. Vol. 23. P.325-331.
Mironenko A.A. The soft-tissue attachment scars in Late Jurassic ammonites from Central Russia // Acta Palaeontologica Polonica. 2015 a. Vol. 60. P.981–1000.
Mironenko A.A. Soft-tissue preservation in the Middle Jurassic ammonite Cadoceras from Central Russia // Swiss Journal of Palaeontology. 2015 b. Vol. 134. no.2. P.281-287
Mironenko A.A. Wrinkle layer and supracephalic attachment area: implications for ammonoid paleobiology // Bulletin of Geosciences. 2015 c. Vol. 90. no.2. P.389-416.
Mironenko A.A. Microstructure of aptychi of Upper Jurassic (Upper Oxfordian) ammonites from Central Russia // Lethaia. 2017 a. in press, DOI 10.1111/let12223.
Mironenko A.A. Siphuncle soft-parts in the Upper Jurassic ammonite Kachpurites fulgens // Palaios. 2017 b. Vol. 32. no.3. P.153-157.
Mironenko A.A., Rogov M.A. First direct evidence of ammonoid ovoviviparity // Lethaia. 2016. Vol. 49. no.2. P.245-260.
Nixon M., Young J.Z. The brains and lives of cephalopods. Oxford: Oxford University Press, 2003. 392 pp.
Norman M.D., Paul D., Finn J., Tregenza T. First encounter with a live male blanket octopus: The world's most sexually size-dimorphic large animal // New Zealand Journal of Marine and Freshwater Research. 2002. Vol. 36. No.4. P.733-736.
Ogura A., Yoshida M., Moritaki T., Okuda Y., Sese J., Shimizu K.K., Sousounis S., Tsonis P. A. Loss of the six3/6 controlling pathways might have resulted in pinhole-eye evolution in Nautilus. // Scientific Reports. 2013. Vol. 3. art. 1432. doi:10.1038/srep01432
Parent H., Westermann G.E.G. Jurassic ammonite aptychi: functions and evolutionary implications // Swiss Journal of Palaeontology. 2016. Vol. 135. no.1. P.101-108.
Polizzotto K., Landman N.H., Klug C. Cameral membranes, pseudosutures, and other soft tissue imprints in ammonoid shells // // In Klug C., Korn D., De Baets K., Kruta I., Mapes R.H. (eds.) Ammonoid paleobiology: from anatomy to ecology. Dordrecht: Springer, 2015. P. 91-109.
Rogov M.A., Mironenko A.A. Patterns of the evolution of aptychi of Middle Jurassic to Early Cretaceous Boreal ammonites // Swiss Journal of Palaeontology. 2016. Vol. 135. no.1. P.139-151.
Shigeno S., Sasaki T., Moritaki T., et al. Evolution of the cephalopod head complex by assembly of multiple molluscan body parts: evidence from Nautilus embryonic development // Journal of Morphology. 2008. Vol. 269. P.1–17
Shigeno S., Takenori S., Boletzky S.V. The origins of cephalopod body plans: a geometrical and developmental basis for the evolution of vertebrate-like organ systems // In: Tanabe K., Shigeta Y., Sasaki T., Hirano H. (eds.) Cephalopods-present and past. Tokyo: Tokai University, 2010. P. 23-34
Stürmer W. Pyrit-Erhaltung von Weichteilen bei devonischen Cephalopoden // Paläontologische Zeitschrift. 1969. Bd. 43. S.10–12.
Tajika A., Naglik C., Morimoto N., Pascual-Cebrian E., Hennhöfer D., Klug C. Empirical 3D-model of the conch of the Middle Jurassic ammonite microconch Normannites, its buoyancy, the physical effects of its mature modifications and speculations on their function. // Historical Biology. 2015. Vol. 27. no.2. P. 181–191.
Tanabe K., Mapes R.H., Sasaki T., Landman N.H. Soft-part anatomy of the siphuncle in Permian prolecanitid ammonoids // Lethaia. 2000. Vol. 33. P. 83–91.
Tanabe, K., Kruta, I., Landman, N.H. Ammonoid buccal mass and jaw apparatus //// In Klug C., Korn D., De Baets K., Kruta I., Mapes R.H. (eds.) Ammonoid paleobiology: from anatomy to ecology. Dordrecht: Springer, 2015 a. P. 439-494.
Tanabe K., Sasaki T., Mapes R.H. Soft-part anatomy of the siphuncle in ammonoids. // // In Klug C., Korn D., De Baets K., Kruta I., Mapes R.H. (eds.) Ammonoid paleobiology: from anatomy to ecology. Dordrecht: Springer, 2015 b. P. 531–544.
Trautschold H. Ueber die Erhaltungszustande russischer ammoniten // Bulletin de la Société Impériale des Naturalistes de Moscou. 1871. T. 43. no.3-4. P. 301–306.
Trueman A.E. The ammonite body-chamber, with special reference to the buoyancy and mode of life of the living ammonite // Quarterly Journal of the Geological Society of London.Q. Journ. Geol. Soc. 1941. Vol. 96. P.339–383.
Ward P. D., Saunders W. B. Allonautilus: a new genus of living nautiloid cephalopod and its bearing on phylogeny of the Nautilida // Journal of Paleontology. 1997. Vol. 71. P.1054-1064.
Westermann G.E.G. Monographie der Otoitidae (Ammonoidea) // Beihefte zum Geologischen Jahrbuch. 1954. Bd. 15. 364 S.
Westermann G.E.G. Ammonoid life and habitat // In Landman, N., Tanabe, K., Davis, R.A. (eds.) Ammonoid Paleobiology. Topics in Geobiology 13. New York: Plenum Press. 1996. P. 606–707.
Zatoń M., Mironenko A.A. Exceptionally preserved Late Jurassic gastropod egg capsules // Palaios. 2015. Vol. 30. P.482-489.
Zeiss A. Weichteile ectocochleater paläozoischer Cephalopoden in Röntgenaufnahmen und ihre paläontologische Bedeutung // Paläontologische Zeitschrift. 1969. Bd. 43. S. 13–27
Ключевые слова (тэги): аммониты, аптихи, аммоноидеи, Wrinkle layer
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 21 сентября 2017 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://ammonit.ru./text/2074.htm#306691">http://ammonit.ru./text/2074.htm#306691</a> |
|
|||||
Ого, какой огромный труд!!! С ума сойти: живородящими были!!!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 21 сентября 2017 года в 13:52:03 |
|||||
 Добавить в избранное
Добавить в избранное 


|
<a href="http://ammonit.ru./text/2074.htm#306711">http://ammonit.ru./text/2074.htm#306711</a> |
|
|||||
Браво Александр! Я в восторге ! В избранное! | ||||||
|
комментарий 1 уровня |
Комментарий создан 21 сентября 2017 года в 18:43:54 |
|||||

|
<a href="http://ammonit.ru./text/2074.htm#306713">http://ammonit.ru./text/2074.htm#306713</a> |
|
|||||
Все так информативно,да еще в одной статье!!!))) Просто здорово! Большое Вам спасибо Александр !! | ||||||
|
комментарий 2 уровня |
Комментарий создан 21 сентября 2017 года в 19:09:12 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#306739">http://ammonit.ru./text/2074.htm#306739</a> |
|
|||||
Александр, большое спасибо!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 21 сентября 2017 года в 22:08:08 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#306811">http://ammonit.ru./text/2074.htm#306811</a> |
|
|||||
Михаил, большое спасибо за этот замечательный вопрос. На что можно ответить сейчас - это на последнюю часть вопроса - про соленость. Я сам в свое время тут цитировал ошибочные версии, но сейчас понятно, что меняется именно соотношение газа и жидкости. То есть имела место именно откачка жидкости, а не изменение ее солености.
| ||||||
|
комментарий 2 уровня |
Комментарий создан 23 сентября 2017 года в 14:54:08 |
|||||

|
<a href="http://ammonit.ru./text/2074.htm#306991">http://ammonit.ru./text/2074.htm#306991</a> |
|
|||||
Значит все-таки газ-жидкость.
| ||||||
|
комментарий 3 уровня |
Комментарий создан 24 сентября 2017 года в 22:22:04 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#306997">http://ammonit.ru./text/2074.htm#306997</a> |
|
|||||
Да, газ-жидкость. Одни авторы пишут, что плохо, когда сифон не достает до жидкости - как ее тогда откачивать? Другие пишут, что плохо, когда он в жидкости, мол тогда он все время должен ее откачивать и тратить энергию, иначе она потечет из него обратно. В общем, гипотез и версий наплодили очень много, все это стоит отдельной и серьезной публикации. | ||||||
|
комментарий 4 уровня |
Комментарий создан 24 сентября 2017 года в 22:39:06 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#306998">http://ammonit.ru./text/2074.htm#306998</a> |
|
|||||
Ну тогда подождем.
| ||||||
|
комментарий 5 уровня |
Комментарий создан 24 сентября 2017 года в 22:52:25 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#306758">http://ammonit.ru./text/2074.htm#306758</a> |
|
|||||
Возьму к руководству. Многое объясняет. Спасибо))) | ||||||
|
комментарий 1 уровня |
Комментарий создан 22 сентября 2017 года в 01:12:43 |
|||||

|
<a href="http://ammonit.ru./text/2074.htm#306795">http://ammonit.ru./text/2074.htm#306795</a> |
|
|||||
Замечательная работа! Узнал много нового про аммонитов.
| ||||||
|
комментарий 1 уровня |
Комментарий создан 23 сентября 2017 года в 09:22:33 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#306796">http://ammonit.ru./text/2074.htm#306796</a> |
|
|||||
Спасибо большое! Это очень познавательно и интересно. | ||||||
|
комментарий 1 уровня |
Комментарий создан 23 сентября 2017 года в 10:59:34 |
|||||

|
<a href="http://ammonit.ru./text/2074.htm#306810">http://ammonit.ru./text/2074.htm#306810</a> |
|
|||||
Большое спасибо за добрые комментарии! Статья стоила времени и сил, но о я очень рад, что это было не зря и что она понравилась! | ||||||
|
комментарий 1 уровня |
Комментарий создан 23 сентября 2017 года в 14:46:26 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#306825">http://ammonit.ru./text/2074.htm#306825</a> |
|
|||||
Как много я не знала :(. Александр, огромное спасибо за столь полезный, интересный и просто обалденный труд :)! Тоже себе в избранное :) | ||||||
|
комментарий 1 уровня |
Комментарий создан 23 сентября 2017 года в 19:00:17 |
|||||

|
<a href="http://ammonit.ru./text/2074.htm#307606">http://ammonit.ru./text/2074.htm#307606</a> |
|
|||||
Редкая публикация! Очень надеюсь, что скоро найдут аммонита вместе с головой! в Ляонине разве нет аммонитов? | ||||||
|
комментарий 1 уровня |
Комментарий создан 1 октября 2017 года в 12:55:34 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#307611">http://ammonit.ru./text/2074.htm#307611</a> |
|
|||||
Я тоже надеюсь! С головой, руками, воронкой и прочими частями тела. А в Ляонине их нет - там континентальные отложения. Там была пресная вода, а головоногие пресную воду принципиально не выносят, это морские животные, обитатели соленых вод. | ||||||
|
комментарий 2 уровня |
Комментарий создан 1 октября 2017 года в 14:02:11 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#307617">http://ammonit.ru./text/2074.htm#307617</a> |
|
|||||
А эти реконструкции меньше актуальны? Автор почему-то изобразил слепого аммонита... http://www.ammonit.ru/text/679.htm | ||||||
|
комментарий 3 уровня |
Комментарий создан 1 октября 2017 года в 14:52:12 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#307618">http://ammonit.ru./text/2074.htm#307618</a> |
|
|||||
Слепого? О какой именно реконструкции речь? Поясните со ссылкой. | ||||||
|
комментарий 4 уровня |
Комментарий создан 1 октября 2017 года в 14:54:57 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#307619">http://ammonit.ru./text/2074.htm#307619</a> |
|
|||||
я отправил тебе сообщение. | ||||||
|
комментарий 5 уровня |
Комментарий создан 1 октября 2017 года в 15:03:09 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#307620">http://ammonit.ru./text/2074.htm#307620</a> |
|
|||||
А мы на "ты" и не переходили. | ||||||
|
комментарий 6 уровня |
Комментарий создан 1 октября 2017 года в 15:08:33 |
|||||
|
<a href="http://ammonit.ru./text/2074.htm#346849">http://ammonit.ru./text/2074.htm#346849</a> |
|
|||||
Саша, нашел статью, что скажешь? https://scienceblogger.livejournal.com/33536.html | ||||||
|
комментарий 1 уровня |
Комментарий создан 6 июля 2019 года в 14:39:45 |
|||||
